This essay explores using the hippocampus as a sequence generator [Buzsáki and Tingley 2018] to precisely time the avoidance action after a failed seek. In essay 31, the animal started with a roaming random search, which turned into a directed seek when it smelled a food odor. If the animal failed to find food after a timeout, it would avoid the area. This essay expands on that model by improving the avoidance action. Previously, the avoidance time was modeled on a cellular timeout, which is imprecise. Instead, we can use an hippocampus sequence to time the avoidance.

This foraging search resembles a Levy walk, which combines a area-restricted brownian walk with longer movements to avoid repeated searches in an area.
In the previous essay, the seek action timed out using astrocyte-managed adenosine in the striatum, but the avoid timeout wasn’t specified, presumably piggybacking on the astrocyte adenosine. Improving the avoidance time and distance can use a sequence generated by the hippocampus. In mice, this kind of distance measurement and timeout is seen in C.pp (posterior parietal cortex) with neurons tiling the delay period, forming a sequence [Harvey et al 2012], [Rajan et al 2016] and in E.ca1 (hippocampus CA1 area) [Pezzulo et al 2017].
Because this essay remains as primitive as possible, and we haven’t added cortex regions yet, we can only add one simplified cortical area. One option is to consider C.pp as a primitive cortex region that can self-generate the necessary sequences. Another option is to consider E.hc (hippocampus) as the main sequence generator and treat C.pp as a later specialization for sophisticated vertebrates like mammals.
The hippocampus can be seen as a sequence generator [Buzsáki and Tingley 2018]. E.hc (hippocampus) sequences are approximately 7s from first place cell neuron to the last neuron in the sequence [Pezzulo et al 2017], and E.hc delay timing is approximately 8s [Abela et al 2015]. Similarly F.pl (prelimbic prefrontal cortex) sequence neurons tile choice encoding for 7s across an experimental trial. C.pp neurons can tile the distance from a start position in mice [Harvey et al 2012], [Rajan et al 2016]. In mice E.ca1 path integration has a maximum of 2m unless extended by landmarks [Fischler-Ruiz et al 2021].
Additionally, the timeout sequence needs a mechanism to trigger it and to drive the avoidance action. I’ll cover a possible trigger from H.sum (supramammillary) and an output from E.hc either directly to H.l (lateral hypothalamus), H.sum, or Poa (preoptic area), or using S.ls (lateral septum as an intermediary.
E.hc place cell for delay
A study in [Fischler-Ruiz et al 2021] studied E.ca1 (CA1 region of hippocampus) with mice traveling a virtual maze, where the liquid reward was 4m from the initial starting point. Instating of waiting for the full 4m, mice expected reward at 2m unless an odor landmark near 2m extended the range to 4m. Neurons in E.ca1 tracked the distance traveled with short time neuron fragment tiling the delay period. A sparse number of neurons active for a short period and as one neuron ends another, new neuron takes its place, like a long thread is made of shorter fibers. This thread frays at around 2m when mice lose track of the task.

Mouse C.pp neurons in virtual maze tile the delay period [Kamiński and Rutishauser 2020]. The above diagram shows the neural tiling. Each row is a single neuron, and time is on the x axis, and the neurons are sorted by their activity. At top is an initial simultaneous burst of many neurons that respond to some event. This large burst is followed by smaller sets of neurons that persist for a small time. When one neuron stops, another neuron takes its place, until the sequence frays and ends.
Roam vs Seek
H.sum is associated with active exploration and is quiet while eating [Kesner et al 2021]. H.sum is activated by food anticipation, food restriction, or gherkin, a hunger peptide [Le May et al 2019].

[Wee et al 2019] H.l (lateral hypothalamus) and Hc (caudal hypothalamus) are both feeding related but are anti-correlated. Hc activates when the zebrafish is hungry and drives the roaming search for food, but when a specific target (paramecium for zebrafish) is detected, H.l activates and Hc deactivates for specific hunting and for eating [Wee et al 2019].
In zebrafish, Hc is comparable to the mammal posterior hypothalamus, which includes H.sum and H.mb (mammillary body), H.tu (tuberal hypothalamus), and H.arc (arcuate nucleus). Zebrafish Hc includes all these areas [Schredelseker and Driver 2020], which are genetically and functionally distinct. Unfortunately this makes it unclear which area is driving the roaming search. H.arc for example is associated with hunger and anti-correlated with eating, using hunger neuropeptides AgRP and NPY in H.arc [Berrios et al 2021]. H.sum is also activated by food anticipation or hunger [Le May et al 2019].
AgRP drops quickly on food cues, drops more slowly for gut nutrient detection, and is slow or permanent in energy balance like blood glucose [Berrios et al 2021]. The specific path for seek dropping AgRP is H.l glutamate to H.dm (dorsal medial hypothalamus) GABA to H.arc AgRP. The H.dm effect disappears with sleep time. RTPP (real-time place preference) is 80% if AgRP is inhibited.
In addition, zebrafish H.l is not identical with the mammal H.l. For example, the orexin and MCH neuropeptide neurons in mammals are part of H.l, while they are in distinct non-H.l areas in zebrafish [Schredelseker and Driver 2020].
In other words, while this essay is taking H.sum as embodying the roam as opposed to H.l embodying the seek, the exact correlations of roam and seek are not yet known.
E.hc input and output
If E.hc drives the seek timeout’s avoid action sustain, how does the seek timeout trigger the E.hc sequence timer to start, and how does E.hc sustain the avoidance? E.hc has three major avoidance action paths it could influence: the obstacle avoidance path through OT (optic tectum) from essay 34, the temporal gradient taxis path through Hb.m – R.ip (medial habenula to interpeduncular nucleus) from essay 33, and the related motivational path through H.l (lateral hypothalamus) and Hb.l (lateral habenula).

The above diagram shows possible input paths to E.hc. On the left, OT (optic tectum) sends visual threat and obstacle information to C.pp and E.por (postrhinal / parahippocampal cortex) via T.lp (lateral posterior / pulvinar thalamus), then to E.mec (medial entorhinal cortex), and finally to E.hc. In the right panel, Hb.m-R.ip sends feedback to E.hc through multiple paths, including H.sum, V.mr (median raphe), R.in (nucleus incepts) and through P.ms (median septum). These two paths differ not only on the information they convey, but also their circuit effects on E.hc. While the left OT path is a data-driven path that excites glutamate neurons, the right Hb.m-R.ip path is a control path that drives E.hc interneuron control circuitry using GABA, ACh (acetylcholine), and 5HT (serotonin) to modulate the action and timing of E.hc.
For this essay, the control path is more interesting, is part because it’s more closely tied to the seek-roam control, and in part because the control circuitry is more fundamental to E.hc operation. The interneuron circuit for the cortex and the hippocampus are highly conserved for all vertebrates, but at some point in evolution they must have been new. Because this essay is exploring adding the first cortical-like area, it needs to address the interneuron controls and not merely assume their existence.

A different, motivational path using V.mr arrives from Hb.l (lateral habenula) instead of the Hb.m – R.ip taxis path. Hb.l is more motivational and Hb.m it more of a physical taxis function. As I’ll be using later in the essay, the full avoid path first travels through the motivational Hb.l, then to E.hc for the avoid time/distance sequence, and finally to Hb.m – R.ip for physical avoidance.

The avoidance output path from E.hc could use one of three action paths. E.hc can drive obstacle-like avoidance through C.pp to OT. Secondly it can drive physical gradient taxis avoidance through Hb.m via P.ts (triangular septum) and P.bac (bed nucleus of the anterior commissure) [Yamaguchi et al 2013]. Finally it can drive motivational avoidance to H.l, H.sum, and Poa (preoptic area) through S.ls (lateral septum). The above diagram shows the obstacle avoidance path through C.pp to OT and the motivational avoidance path through H.l, H.sum, and Poa. (Note: Poa to M.pag to MLR is more direct avoidance than motivational.) The Poa path drives movement directly through M.pag (periaqueductal gray) and MLR (midbrain locomotor region), which the diagram omits for brevity.

Another E.hc motivational output uses S.v (ventral striatum / nucleus accumbens) and P.v (ventral pallidum / endopeduncular nucleus) to H.l and Vta (ventral tegmental area) for seeking and to Hb. for avoidance. Although this path is similar to the S.ls motivational path above, in mammals at least it’s a distinct circuit. Unlike the S.ls path, the S.v motivational path does not target H.sum or Poa but does strongly target Hb. Note that S.v and S.ls are similar structures both derived from the same progenitor domain LGE (lateral ganglionic eminence) and is neighbors with S.v, also known as nucleus accumbens septi, Latin for nucleus adjacent to the septum.

Mammals also have a direct E.hc output to Hb.m-R.ip via P.ts (triangular septum) and P.bac (bed nucleus of the anterior commissure, which is sometimes identified with part of P.bst bed nucleus of the stria terminalis) [Proulx et al 2014]. As described in previous essays, Hb.m has a direct function for phototaxis [Chen and Engert 2014], chemotaxis [Beretta et al 2012], thermotaxis, and some social conflict [Agetsuma et al 2010]. This mammal P.ts/P.bac is understudied, or I haven’t found any study describing its function, but by its connection E.hc could drive taxis via the Hb.m-R.ip connection. This connection does not exist in lamprey [Stephenson-Jones et al 2012], but I haven’t read any studies on this connectivity for other non-mammal animals.
Ancestral vertebrate forebrain
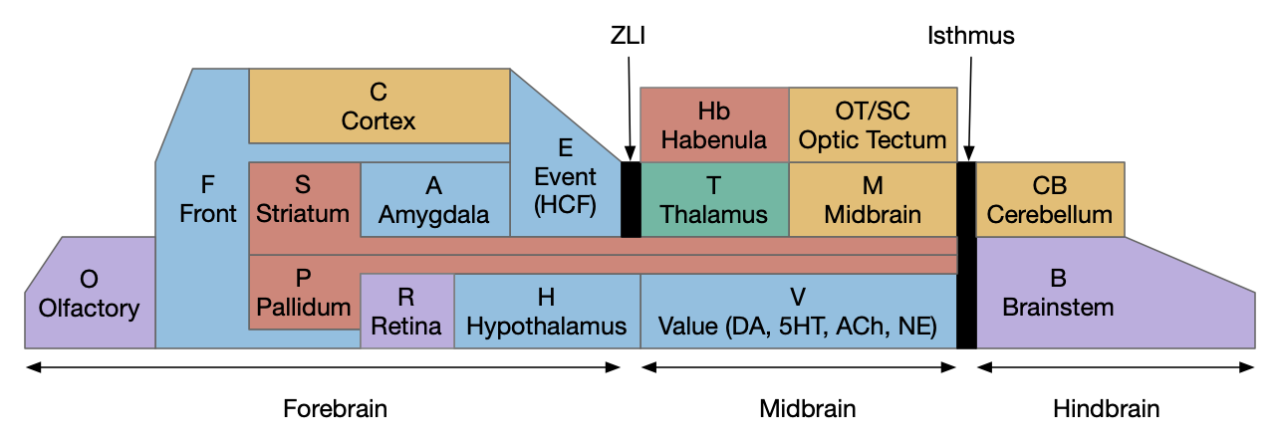
Stepping back to cover the ancestral vertebrate forebrain (cortex and basal ganglia) to understand likely primitive areas, particularly the E.hc equivalent. Ancestral models generally use Pa (pallium) to describe cortical-like areas and divide pallial areas into multiple regions, generally from four to six, depending on the theory. These areas are named by their location, where MPa (medial pallium) is in the middle, DPa (dorsal pallium) is on top, and LPa (lateral pallium) on the side.

The above diagram shows the quadripartite model of the ancestral vertebrate forebrain [Hegarty et al 2024], [Pessoa et al 2019]. The dorsal areas are cortical-like and the ventral (basal) areas are striatal and pallidal (basal ganglia). The four areas match most vertebrate structure. MPa (medial pallium) is hippocampal, DPa (dorsal pallium) is sensory neocortex, LPa (lateral pallium) is the insula cortex (eating and tasting), E.lec (lateral entorhinal) and F.ofc (orbitofrontal), and VPa (ventral pallium) is the olfactory cortex and amygdala. In the diagram, the notch at the top is deliberate because the neural tube is formed by curling up a neural sheet until the two ends nearly match at the top. In most vertebrates the two ends at MPa curl more until they reach the base, forming two hemispheres, but in teleost fish the two ends curl out, putting MPa on lateral outside (Dl in teleosts) and VPa in the middle (Dm in teleosts) [Hegarty et al 2024], [Roth and Dicke 2013], [Porter and Mueller 2020].
Although this model is useful, it does minimize differences between vertebrates. For example, the mammalian DPa, the “neocortex,” doesn’t nicely match up with reptiles, instead the amygdala-like areas (DVR) take a larger role.

In amniotes (lizards, birds, and mammals) E.hc has a similar structure, including E.dg (dentate gyrus), E.ca1, E.ca3, E.sub (subiculum), E.lec (lateral entorhinal cortex), and E.mec (medial entorhinal cortex) [Medina et al 2017]. As mentioned previously, although the fish Dl is likely an E.hc-like structure, its internals differ from the amniotes. If the fish Dl was a subset of the mammal E.hc, it would be useful to know which areas are more primitive.
The lamprey is interesting because it may not have an E.hc / MPa equivalent at all. A recent genetic cell analysis [Lamanna et al 2023], [Hervas-Sotomayor 2023] suggests that MPa is not ancestral because the medial pallial area in lampreys matches H.em (prethalamic eminence) instead, which is more closely associated with the hypothalamus and habenula, not the forebrain and specifically not matching hippocampal markers. In addition, the lamprey extended amygdala is distinct, well-defined, and separate from the lateral pallium, suggesting it may be more helpful to treat the extended amygdala as a distinct area instead of as part of the pallium. In the case of mammals, that distinction may not apply to A.bl (basolateral amygdala), which resembles the cortical DPa more than other parts of the amygdala [Moreno and González 2007]. [Lammana et al 2023] suggest that the quadripartite model may only apply to later vertebrates, and lampreys having an undifferentiated pallium, an amygdala, and a H.em with unknown purpose. These genetic results reinforce earlier suggestions that the lamprey “MPa” is actually an expended H.em.
For this study, which only uses the hippocampus / MPa, the lack of a lamprey MPa suggests that the sequential hippocampal model is a later vertebrate development. Even if accurate, it would be unknown if it developed specifically with jawed vertebrates (gnathostomes) or if it was an innovation ozone of the many preceding jawless vertebrates.
C.pp navigation delay
In mice, C.pp (posterior parietal cortex) tracks time and distance from a start position with neurons that tile the delay period, as shown in the earlier neuron firing diagram. Each neuron is only active for a fragment of the task, and as one neuron leaves the ensemble, another starts [Harvey et al 2012]. In their task, C.pp was required for memory related success, not needed for cue seeking.
C.pp is “vision for action” [Goodale and Milner 1992]. C.pp tuned to referred self motion and acceleration [Whitlock et al 2012]. In this context, the timing we need it motion timing, so self motion is critical. Some C.pp neurons fire up to 500ms before action and drop immediately on action.
Although these locomotion delays match what the essay needs, Cpp has a number of problems for this purpose. First, C.pp is part of the neocortex, which is specific to mammals. Birds, reptiles, and fish have a cortex (called pallium), but the area around C.pp is organized differently with a larger amygdala area (DVR) than cortical (dorsal pallium) [Aboitiz et al 2003]. Mammalian E.hc as an odor-motor region with 1D maps [Aboitiz and Montiel 2015].
Second, the most direct subcortical C.pp output is to OT (optic tectum), but otherwise C.pp requires C.mo (motor cortex) for any action. But in amphibians, the OT does not receive input from a cortical equivalent, but connects directly with the striatum [Pessoa et al 2019]. In the context of the essay, the OT doesn’t seem related to the place avoidance needed.
Third, the C.pp inputs are primarily indirect through other cortical areas, although using T.lp (lateral posterior thalamus) from OT. So, if C.pp is a primitive cortical area, it seems much more likely to be an OT-focused area, not a foraging area.
As a counterpoint, the electric fish Dc also has direct output to OT, which suggests a C.pp-like role. Dc is driven by the Dl (lateral cortex, possible E.dg) through DD (dorsal cortex, as possible E.ca3) [Fotowat et al 2019]. That study suggested the entire Pa (cortex) area is silent unless actively sensing and sequences were not studied. Other research suggests that only Dl.v (ventral Dl) is equivalent to E.hc, particularly to E.sub [Hegarty et al 2024]. [Rodríguez-Expósito et al 2017], which makes the comparison more tenuous, unless DD is something like C.rs (retrospenial cortex), which in mammals is between E.hc and C.pp.
Similarly, the lamprey LPA (lateral pallium / cortex) has a direct OT projection [Suryanarayana et al 2022], giving it a possible C.pp-like role, but the lamprey does not appear to have a hippocampus equivalent [Lamanna et al 2023]. It does have an expanded H.em (prethalamic eminence) which is physically located where the MPa (medial pallium / hippocampus) would be. Unfortunately, no current studies have covered H.em function in the lamprey, but it seems likely to be significantly different from the hippocampus.
In favor of C.pp as relevant here, a study of electric fish shows a path from Dl (E.dg – dentate gyrus equivalent) to Ddi (E.ca3 equivalent) to Dc (C.rs.5/C.pp.5 equivalent) to OT [Fotowat et al 2019]. However, in that study the entire Pa is silent except when swimming or active sensing (electro-sensing in this case), and it’s unclear if there are any sequences as in the mouse C.pp.
A different description of Dl points out that it primarily projects to Poa and Hc (caudal hypothalamus) [Northcutt 2006]. The fish Hc .sum set of efferents is that Dl projects to the area that includes H.arc, H.mb, H.sum, H.pm, and H.tu, which are developmentally conserved between fish and mammals [Wullimann 2022]. Because these areas have different functions, it would be useful to known exactly which specific areas Dl projects to. In mammals, Poa and H.sum are interconnected and are both associated with exploration and roaming [Escobedo et al 2023], [Ryoo et al 2021].
H.sum
As mentioned previously, H.sum is part of the basal hypothalamus and is highly conserved, existing in sharks [Santos-Durán et al 2022], teleost fish [Wullimann 2022], amphibians and reptiles [Domínguez et al 2016], birds [Kim DW et al 2022], and mammals [Bedont et al 2015], [Croizier et al 2015], [Ferran et al 2015]. I haven’t found an equivalent lamprey hypothalamus study, which would be especially interesting because of the lamprey’s lack of MPa / E.hc equivalent. If H.sum does exist in lampreys, its function and connectivity might show a simpler, more primitive function before adding MPa / E.hc capabilities.
H.sum has several distinct circuits, some with overlapping behavior. H.sum tac1 (Substance-P marker) is highly associated with voluntary locomotion, but not with E.hc theta [Farrell et al 2021]. H.sum is associated with active exploration and produces RTPP (real-time place preference) but is quiet while eating [Kesner et al 2021]. H.sum to Poa (preoptic area) projections are associated with avoidance to threads and shows a strong RTPA (real-time place avoidance) but not anxiety or CPA (conditioned place avoidance) [Escobedo et al 2023]. The H.sum to Poa connection has collaterals to E.ca2 but not to Hb.m or E.dg (hippocampus dentate gyrus). H.sum stimulation inhibits eating and its inhibition is required for eating. The projections to E.dg are related to object novelty and open field exploration [Pan et al 2004], [Chen S et al 2020], while the projection to E.ca2 is related to social novelty and temporal memory [Chen Z et al 2022], [Thirtamara et al 2024]. H.sum is also part of the food reward circuit and is reinforcing [Ikemoto 2010].
E.hc – hippocampus as sequence generator
One model of E.hc (hippocampus) is as a general sequence generator [Buzsáki and Tingley 2018]. The idea notes that E.hc is blind to its inputs, whether olfactory, visual, or vestibular, but what is consistent it its ability to tile gaps between events, and its internal timing structures such as fitting gamma (40-100Hz) bursts inside theta cycles (~8Hz), such as a consistent seven-ish game cycle bursts within a single theta cycle.
E.ca2 – hippocampal CA2
E.hc is a complicated structure with many parts, but must have evolved from an initial core area. Studies with other vertebrates show comparable areas to the hippocampus E.dg (dentate gyrus), E.ca1 (CA1 area), E.ca3 (CA3 area), and E.sub (subiculum area).
For this essay, I’m considering E.ca2 as the most primitive because it’s strongly tied to sequence generation [Bhasin and Nair 2022], [He et al 2021], [Lehr et al 2021], [MacDonald and Tonegawa 2021], [Stöber et al 2020] and is reciprocally connected with H.sum, which I’m already using for its connection with RTPA and exploration. E.ca2 is reciprocally connected with H.sum in the early embryonic state, earlier than E.dg and E.ca3 connections [Diethorn and Gould 2023].
In mice, E.ca2 sustains internal sequence memory during delay [Lehr and Stöber 2021]. When E.ca2 is disabled, sequences are destabilized in E.ca1 [Lehr et al 2021]. Interestingly, during sleep E.ca2 appears to fire consistently to remember the animal’s current location [Kay et al 2016].
E.ca2 is distinguished from E.ca3 by its lack of E.dg input [Insausti et al 2023], making it an interesting candidate for an early area because it has fewer dependencies and requirements.
Neural models for sequences and persistence
Neural models for sequences are much less studied than models for persistence and memory. I’m using sequences for a delay period and delay periods are well studied. In behaviorist experiments, “trace conditioning” has a short delay between the cue to be remembered and the food reward or shock punishment.
Before approaching sequences, it seems best to consider persistence, which is better studied, such as persistent working memory. [Zylberberg and Strowbridge 2017] reviews multiple models of persistence. The main difference I’ll highlight is between models focused on intrinsic cellular responses such as bursting, in contrast with models that focus on recurrent connectivity, such as Hopfield networks [Hopfield 1982]. Experimentally, the evidence between cell-autonomous vs recurrent network is an open question [Kamiński and Rutishauser 2020].
Persistent bursts are 3-6 spikes [Zylberberg and Strowbridge 2017]. Epilepsy is related to these bursts and may be a failure in inhibitory circuits to contain expansion of these bursts. These bursts are enabled by metabotropic receptors such as mACh (acetylcholine metabotropic receptor). The hindbrain includes bursting neurons, such as VOR (vestibuloocular reflex), long plateau potentials in spinal motoneurons with Ca2+ (calcium) channels and with K+ (potassium) channels. These bursting mechanisms can rebound after inhibition.
Recurrent models of persistence
Recurrent models of persistence use feedback, recurrent connections of neurons back to the same area. If properly calibrated, these loops produce produce attractors that can remember data [Hopfield 1982]. A difficulty in recurrent models is the issue of fast neural circuits (10-20ms) extending to seconds, and how to maintain stability when 100-fold timescales are required [Zylberberg and Strowbridge 2017]. One common response is to use longer time circuits such as NMDA receptors with 100ms time constants. The longer basal constants reduce the stability issues. For example, one of the models for recurrent storage uses NMDA in a model of the VOR (vestibular-ocular reflex) [Seung 1996], where the eye target is maintained in a line attractor.
Criticisms of recurrence and attractors note the requirement for fine tuning [Zylberberg and Strowbridge 2017]. Continuous attractors require fine turning. For example, line attractors require precise recurrence, while noise and modulation affects the tractors. In addition, the timescale of excitation vs inhibition is important to the stability. In addition neurons tend to switch abruptly between discrete states as opposed to smooth, continuous variation. To reduce the issue, finely spaced concrete attractors are more tolerant than continuous attractors.
Continuous attractors use a smooth neuron rate encoded values, but firing rate models difficult to match actual neuron behavior [Compte et al 2000]. Spontaneous activity at 3.5Hz does not trigger attractor. [Lundqvist et al 2016] notes that individual neurons bridging a multi-second memory delay is rare, but a simple attractor model would have multiple neurons bridging the entire gap. [Cui and Strowbridge 2018] also note there is little experimental support for recurrent synaptic reverberation.
Intrinsic neural persistence
Intrinsic neural persistence models focuses more on cellular mechanisms for persistence, where an individual neuron fires for longer times, such as 2s without requiring external prompting like recurrence. Since the Lundqvist criticism [Lundqvist et al 2016] also applies here, that single neurons do not cover the entire gap, intrinsic cellular persistence alone is insufficient. However, extending the time constant from the 100ms of NMDA to 500ms or even multiple seconds reduces the need for precise calibration.
Persistent firing exists in C (cortex) and Ob (olfactory bulb) using cellular mechanisms, initiates by a burst of AP (action potentials) [Cui and Strowbridge 2018]. Many cellular persistent firings are enabled by m1.q (acetylcholine metabotropic receptor tied to G-q/11 proteins), and additionally require cellular Ca2+ (calcium ions) for bursting, triggered by the input bursts. ACh enhances excitability by reducing ERG (ether-a-go-go) K+ (potassium currents). The ERG K+ currents typically suppress bursting and persistence. By suppressing ERG K+, the ACh to m1.q path disinhibits bursting and persistence. ERG is highly expressed in deep C (layer 5/6 cortex), Snc DA (substantia nigra compacta dopamine), hindbrain, and E.ca1. ERG blockers abolish persistent firing in C.tea (temporal association area) and F.pfc (prefrontal cortex).
In the Cui and Strowbridge study [Cui and Strowbridge 2018], ACh enables persistence for at least 5s at 4.3Hz to 6.4Hz. With normal, non-ACh ERG behavior, input bursts produce a short AP burst followed by suppressed output. When ACh blocks ERG currents, input bursts produce an AP burst followed by persistent firing that requires Ca2+. In short, ACh enables persistent firing in the range of 500ms to 2ms for a single neuron without needing external recurrence.
Other studies show similar modulated persistence in other areas. In E.sub, ACh stimulation of m1.q enables extended plateau potentials [Kawasaki et al 1999], allowing sustained tonic firing. In layer 5 cortex, when ACh enables persistent firing, even the smallest apical depolarization produces L5 repetitive bursts [Schwindt and Crill 1999]. The plateau potential and prolonged response is over 400ms. Similar effects for ACh and m1.q promote sustained persistence in CB (cerebellum) and O.mt (olfactory bulb mitral cell) [Bauer and Schwarz 2018]
Tiling and threading
Combining longer persistent intrinsic cellular activity with recurrence provides many options for persistence and sequence generation. As mentioned above, mouse C.pp neurons in a virtual maze tiles a delay period [Kamiński and Rutishauser 2020]. Each neuron has an intrinsic persistent between 500ms and 1500ms, and only a sparse number like 10% are active at a time [Pastalkova et al 2008]. As one neuron ends its extended 500ms activity, another neuron takes its place. When the animal acts, the sequence is extinguished [Zylberberg and Strowbridge 2017].

One component of the sequence control is the E/I (excitation / inhibition) balance, managed by cortical interneurons. The E / I balance prevents runaway excitation, as in epilepsy, and instead restricts activity to a small subset of neurons. This balance is often oscillatory and combined with winner-take-all systems, where the first neurons to fire suppress slower neurons. In the case of sequences, when one neuron stops firing when its internal persistence ends, the E / I balance shifts towards allowing a new neuron to take its place.

An older model of sequences is the synfire chain, where each neuron triggers the next neuron in the sequence. An argument against the strict synfire chain its developmental plausibility, because it requires precise connectivity.

Instead of a precise synfire chain or the precise dynamics required for recurrent systems like [Hopfield 1982], [Rajan et al 2016] proposes a developmental model for sequences. Consider a set of neurons randomly connected, and modify only some of the connections to produce sequences. Neuron development includes calibration and pruning during early development, including intrinsic firing that would allow for sequence selection. Once the network is formed, sequences propagate by cooperation between recurrence and the input.

Because sequences are dynamic, their output may require a projection layer to a more meaningful subspace [Kamiński and Rutishauser 2020]. For example, this essay only needs a single active signal to continually drive the avoidance until the sequence ends.

The diagram above shows how these ideas fit together. The pyramidal neurons in the middle form the backbone of the sequence. Each neuron has an intrinsic persistent firing for ~500ms. The PV (parvalbumin) interneurons prevent more than one neuron firing. Because the current neuron is already firing, it blocks the next neuron in the chain until the first neuron completes. The sequence is decoded by a separate output layer, such as S.ls (lateral septum) or E.sub (subiculum) before sending to the destination.
Simulation choices
Taking the above research into consideration, the simulation needs to make some choices of a plausible action path. Because this essay builds on the previous essay 31, which used S.v to Hb.l as a seek timeout, it makes sense to continue that path from Hb.l to P.ms and E.hc via V.mr and H.sum.

The output side has two tempting directions: E.hc to Poa and H.sum via S.ls, or E.hc to Hb.m-R.ip via S.ts and S.bac. The E.hc to Hb.m-R.ip has an advantage of exploring differences between Hb.l and Hb.m. One model is Hb.l as the motivational avoid and Hb.m as the physical taxis avoid, where evolution secondarily adds motivational Hb.l avoid as a more sophisticated indirection to the older Hb.m direct avoid taxis. The other option using S.ls to Poa and H.sum has the advantage of possibly being more primitive, because the P.ts / P.bac to Hb.m path may be restricted to mammals and therefore newer. The S.ls path also has the advantage of introducing the strong E.hc to hypothalamus connection via S.ls.
Sequence design
Because the simulation is a toy model where understandability is important, the simulation doesn’t directly implement individual neurons. One advantage is that neuron firing is generally sparse. For example, only about 10% of O.pir (olfactory cortex) neurons fire for any particular odor. Taking an idea from high-dimensional computing [Laiho et al 2015], an engram (neural assembly) can be represented by a vector of digits. Each digit is essentially one sparse neuron selected from a larger set. Base-16 is one neuron selected from 16. This digital representation can be displayed as a normal hex number such as 78af. For display purposes, base-64 is the largest feasible base, because its numerical digits plus lower and upper letters produce 62 with two special digits. Zero is reserved to represent no neurons firing in the digit. So 4305 represents 3 neurons firing, one from each of three active digits with the last digit silent.
In natural sequences, after one neuron falls out, a new random neuron takes its place. In simulation, this randomness adds complexity because to avoid repetition it would need to keep track of earlier values. For neurons this repetition avoidance is built-in because neurons can have a refractory period where they’re less responsive, giving fresh neurons a better chance to be chosen. Instead, the simulation reserves the lowest bits of each digit for sequences, leaving the highest bits for the random engram. The sequence also updates each digit sequentially instead of randomly. Zero represents the end of the sequence when all neurons stop firing. A sequence with a single base-16 digit might look like “8, 9, a, b, 0” or “4, 5, 6, 7, 0” if two bits are sequence bits, and a sequence with two digits might look like “84, 85, 95, 96, a6, a7, ....“
Simulation results
The following diagram shows a sample trajectory where the animal first seeks to the center of the odor plume and spins at the center until the S.v striatum adenosine times out as in essay 31. The timeout triggers a motivational avoid, representing V.mr and H.sum via Hb.l. The motivational avoid starts the E.hc sequence via P.ms, which activates the physical avoid representing either S.ls to Poa or Hb.m to R.ip depending on interpretation. When the sequence ends, E.hc stops driving the physical avoid, and the animal resumes its roaming search.

Discussion
This essay raises a number of issues, mainly because the situation is understudied, and because it involves many regions that are generally studied independently. As a reminder, this essay is at best a thought experiment.
The lamprey’s lack of a MPa / E.hc (medial pallium / hippocampus) is the first interesting issue. H.em (prethalamic eminence) in the expected MPa location is also interesting. What does H.em do? In mammals it’s a progenitor area that disappears, and it produces neurons for P.epn (endopeduncular nucleus) and Cajal-Retzius neurons that populate neocortical layer 1 and organize the neocortical layers. But in lampreys the H.em functionality may be entirely different. It its function an any way similar to a primitive E.hc? Does the lamprey lack all internally generated sequences?
In addition, what happens in lampreys to all the areas connected with MPa / E.hc? Does H.sum and H.mb (mammillary body) exist in lampreys, and if it exists, what does it do? Similar questions for V.mr R.nin (nucleus incepts) and S.ls and P.ms. If E.hc is missing in lampreys, studying those areas might give more information about their function.
More Zebrafish studies about the Dl / E.hc would also be very interesting, because it seems to have a different structure from the common amniote E.hc. Consider the hunger study with the reciprocal activation of Hc (caudal hypothalamus) and H.l (lateral hypothalamus) when the animal detects a food cue [Wee et al 2019]. Because Hc is composed of H.sum, H.mb, H.tu and H.arc, which have distinct functions, learning exactly which areas are activated by hunger and suppressed during seek would be interesting. H.sum and H.mb are also understudied in zebrafish. Do they have similar connectivity to Dl (MPa / E.hc) and function as in mammals? Does Dl have similar sequential functionality? It does have SWF (sharp wave ripples) [Blanco et al 2024], which suggests zebrafish has Dl sequences, but does it have the same threading / tiling of delays?
Although this essay has used E.ca2 as the center of E.hc sequences, but E.ca2 has only been identified in mammals. Genetic transcription comparisons with other animals suggests E.ca3 as better conserved. If E.ca2 is a mammalian innovation, how does that affect how E.hc works in non-mammals?
References
Abela AR, Duan Y, Chudasama Y. Hippocampal interplay with the nucleus accumbens is critical for decisions about time. Eur J Neurosci. 2015 Sep;42(5):2224-33.
Aboitiz F, Morales D, Montiel J. The evolutionary origin of the mammalian isocortex: towards an integrated developmental and functional approach. Behav Brain Sci. 2003 Oct;26(5):535-52; discussion 552-85.
Aboitiz F, Montiel JF. Olfaction, navigation, and the origin of isocortex. Front Neurosci. 2015 Oct 27;9:402.
Agetsuma M., Aizawa H., Aoki T., Nakayama R., M. Takahoko, M. Goto, T. Sassa, R. Amo, T. Shiraki, K. Kawakami, et al. The habenula is crucial for experience-dependent modification of fear responses in zebrafish Nat. Neurosci., 13 (2010), pp. 1354-1356
Bauer CK, Schwarz JR. Ether-à-go-go K+ channels: effective modulators of neuronal excitability. J Physiol. 2018 Mar 1;596(5):769-783.
Bedont JL, Newman EA, Blackshaw S. Patterning, specification, and differentiation in the developing hypothalamus. Wiley Interdiscip Rev Dev Biol. 2015 Sep-Oct;4(5):445-68.
Beretta CA, Dross N, Guiterrez-Triana JA, Ryu S, Carl M. Habenula circuit development: past, present, and future. Front Neurosci. 2012 Apr 23;6:51.
Berrios J, Li C, Madara JC, Garfield AS, Steger JS, Krashes MJ, Lowell BB. Food cue regulation of AGRP hunger neurons guides learning. Nature. 2021 Jul;595(7869):695-700.
Bhasin G, Nair IR. Dynamic Hippocampal CA2 Responses to Contextual Spatial Novelty. Front Syst Neurosci. 2022 Aug 8;16:923911.
Buzsáki G, Tingley D. Space and Time: The Hippocampus as a Sequence Generator. Trends Cogn Sci. 2018 Oct;22(10):853-869.
Chen S, He L, Huang AJY, Boehringer R, Robert V, Wintzer ME, Polygalov D, Weitemier AZ, Tao Y, Gu M, Middleton SJ, Namiki K, Hama H, Therreau L, Chevaleyre V, Hioki H, Miyawaki A, Piskorowski RA, McHugh TJ. A hypothalamic novelty signal modulates hippocampal memory. Nature. 2020 Oct;586(7828):270-274.
Chen X, Engert F. Navigational strategies underlying phototaxis in larval zebrafish. Front Syst Neurosci. 2014 Mar 25;8:39.
Chen Z, Chen G, Zhong J, Jiang S, Lai S, Xu H, Deng X, Li F, Lu S, Zhou K, Li C, Liu Z, Zhang X, Zhu Y. A circuit from lateral septum neurotensin neurons to tuberal nucleus controls hedonic feeding. Mol Psychiatry. 2022 Dec;27(12):4843-4860.
Compte A, Brunel N, Goldman-Rakic PS, Wang XJ. Synaptic mechanisms and network dynamics underlying spatial working memory in a cortical network model. Cereb Cortex. 2000 Sep;10(9):910-23.
Croizier S, Chometton S, Fellmann D, Risold PY. Characterization of a mammalian prosencephalic functional plan. Front Neuroanat. 2015 Jan 6;8:161.
Cui ED, Strowbridge BW. Modulation of Ether-à-Go-Go Related Gene (ERG) Current Governs Intrinsic Persistent Activity in Rodent Neocortical Pyramidal Cells. J Neurosci. 2018 Jan 10;38(2):423-440.
Diethorn EJ, Gould E. Postnatal development of hippocampal CA2 structure and function during the emergence of social recognition of peers. Hippocampus. 2023 Mar;33(3):208-222.
Domínguez L, González A, Moreno N. Patterns of hypothalamic regionalization in amphibians and reptiles: common traits revealed by a genoarchitectonic approach. Front Neuroanat. 2015 Feb 3;9:3.
Escobedo Abraham, Holloway Salli-Ann, Votoupal Megan, Cone Aaron L, Skelton Hannah E, Legaria Alex A., Ndiokho Imeh, Floyd Tasheia, Kravitz Alexxai V., Bruchas Michael R., Norris Aaron J. (2023) Glutamatergic Supramammillary Nucleus Neurons Respond to Threatening Stressors and Promote Active Coping eLife 12:RP90972
Farrell JS, Lovett-Barron M, Klein PM, Sparks FT, Gschwind T, Ortiz AL, Ahanonu B, Bradbury S, Terada S, Oijala M, Hwaun E, Dudok B, Szabo G, Schnitzer MJ, Deisseroth K, Losonczy A, Soltesz I. Supramammillary regulation of locomotion and hippocampal activity. Science. 2021 Dec 17;374(6574):1492-1496.
Ferran JL, Puelles L, Rubenstein JL. Molecular codes defining rostrocaudal domains in the embryonic mouse hypothalamus. Front Neuroanat. 2015 Apr 17;9:46.
Fischler-Ruiz, W., Clark, D. G., Joshi, N. R., Devi-Chou, V., Kitch, L., Schnitzer, M., … & Axel, R. (2021). Olfactory landmarks and path integration converge to form a cognitive spatial map. Neuron, 109(24), 4036-4049.
Fotowat H, Lee C, Jun JJ, Maler L. Neural activity in a hippocampus-like region of the teleost pallium is associated with active sensing and navigation. Elife. 2019 Apr 3;8:e44119.
Goodale MA, Milner AD. Separate visual pathways for perception and action. Trends Neurosci. 1992 Jan;15(1):20-5.
Harvey CD, Coen P, Tank DW. Choice-specific sequences in parietal cortex during a virtual-navigation decision task. Nature. 2012 Mar 14;484(7392):62-8.
He H, Boehringer R, Huang AJY, Overton ETN, Polygalov D, Okanoya K, McHugh TJ. CA2 inhibition reduces the precision of hippocampal assembly reactivation. Neuron. 2021 Nov 17;109(22):3674-3687.e7.
Hegarty BE, Gruenhagen GW, Johnson ZV, Baker CM, Streelman JT. Spatially resolved cell atlas of the teleost telencephalon and deep homology of the vertebrate forebrain. Commun Biol. 2024 May 21;7(1):612.
Hervas Sotomayor, F. (2024). Tracing the origins and evolution of the vertebrate brain: a comparative cellular study in anamniotes (Doctoral dissertation).
Hopfield JJ. Neural networks and physical systems with emergent collective computational abilities. PNAS. 1982;79(8):2554–58.
Ikemoto S. Brain reward circuitry beyond the mesolimbic dopamine system: a neurobiological theory. Neurosci Biobehav Rev. 2010 Nov;35(2):129-50.
Insausti, R., Muñoz‐López, M., & Insausti, A. M. (2023). The CA2 hippocampal subfield in humans: A review. Hippocampus, 33(6), 712-729.
Kamiński J, Rutishauser U. Between persistently active and activity-silent frameworks: novel vistas on the cellular basis of working memory. Ann N Y Acad Sci. 2020 Mar;1464(1):64-75.
Kawasaki H, Palmieri C, Avoli M. Muscarinic receptor activation induces depolarizing plateau potentials in bursting neurons of the rat subiculum. J Neurophysiol. 1999 Nov;82(5):2590-601.
Kay K, Sosa M, Chung JE, Karlsson MP, Larkin MC, Frank LM. A hippocampal network for spatial coding during immobility and sleep. Nature. 2016 Mar 10;531(7593):185-90.
Kesner AJ, Shin R, Calva CB, Don RF, Junn S, Potter CT, Ramsey LA, Abou-Elnaga AF, Cover CG, Wang DV, Lu H, Yang Y, Ikemoto S. Supramammillary neurons projecting to the septum regulate dopamine and motivation for environmental interaction in mice. Nat Commun. 2021 May 14;12(1):2811.
Kim DW, Place E, Chinnaiya K, Manning E, Sun C, Dai W, Groves I, Ohyama K, Burbridge S, Placzek M, Blackshaw S. Single-cell analysis of early chick hypothalamic development reveals that hypothalamic cells are induced from prethalamic-like progenitors. Cell Rep. 2022 Jan 18;38(3):110251.
Lamanna F, Hervas-Sotomayor F, Oel AP, Jandzik D, Sobrido-Cameán D, Santos-Durán GN, Martik ML, Stundl J, Green SA, Brüning T, Mößinger K, Schmidt J, Schneider C, Sepp M, Murat F, Smith JJ, Bronner ME, Rodicio MC, Barreiro-Iglesias A, Medeiros DM, Arendt D, Kaessmann H. A lamprey neural cell type atlas illuminates the origins of the vertebrate brain. Nat Ecol Evol. 2023 Oct;7(10):1714-1728.
Le May MV, Hume C, Sabatier N, Schéle E, Bake T, Bergström U, Menzies J, Dickson SL. Activation of the rat hypothalamic supramammillary nucleus by food anticipation, food restriction or ghrelin administration. J Neuroendocrinol. 2019 Jul;31(7):e12676.
Lehr AB, Kumar A, Tetzlaff C, Hafting T, Fyhn M, Stöber TM. CA2 beyond social memory: Evidence for a fundamental role in hippocampal information processing. Neurosci Biobehav Rev. 2021 Jul;126:398-412.
Lehr AB, Stöber TM. Differential involvement of CA2 in internally vs. externally driven hippocampal sequences. Proc Natl Acad Sci U S A. 2021 Sep 21;118(38):e2110671118.
Lundqvist M, Rose J, Herman P, Brincat SL, Buschman TJ, Miller EK. Gamma and Beta Bursts Underlie Working Memory. Neuron. 2016 Apr 6;90(1):152-164.
MacDonald CJ, Tonegawa S. Crucial role for CA2 inputs in the sequential organization of CA1 time cells supporting memory. Proc Natl Acad Sci U S A. 2021 Jan 19;118(3):e2020698118.
Medina L, Abellán A, Desfilis E. Contribution of Genoarchitecture to Understanding Hippocampal Evolution and Development. Brain Behav Evol. 2017;90(1):25-40.
Moreno N, González A. Evolution of the amygdaloid complex in vertebrates, with special reference to the anamnio-amniotic transition. J Anat. 2007 Aug;211(2):151-63.
Northcutt, R. G. (2006). Connections of the lateral and medial divisions of the goldfish telencephalic pallium. Journal of Comparative Neurology, 494(6), 903-943.
Pan, Wei-Xing, and Neil McNaughton. The supramammillary area: its organization, functions and relationship to the hippocampus. Progress in neurobiology 74.3 (2004): 127-166.
Pastalkova E, Itskov V, Amarasingham A, Buzsáki G. Internally generated cell assembly sequences in the rat hippocampus. Science. 2008 Sep 5;321(5894):1322-7.
Pessoa L, Medina L, Hof PR, Desfilis E. Neural architecture of the vertebrate brain: implications for the interaction between emotion and cognition. Neurosci Biobehav Rev. 2019 Dec;107:296-312.
Pezzulo G, Kemere C, van der Meer MAA. Internally generated hippocampal sequences as a vantage point to probe future-oriented cognition. Ann N Y Acad Sci. 2017 May;1396(1):144-165.
Porter BA, Mueller T. The Zebrafish Amygdaloid Complex – Functional Ground Plan, Molecular Delineation, and Everted Topology. Front Neurosci. 2020 Jul 16;14:608.
Proulx CD, Hikosaka O, Malinow R. Reward processing by the lateral habenula in normal and depressive behaviors. Nat Neurosci. 2014 Sep
Rajan K, Harvey CD, Tank DW. Recurrent Network Models of Sequence Generation and Memory. Neuron. 2016 Apr 6;90(1):128-42.
Rodríguez-Expósito, B., Gómez, A., Martín-Monzón, I., Reiriz, M., Rodríguez, F., & Salas, C. (2017). Goldfish hippocampal pallium is essential to associate temporally discontiguous events. Neurobiology of Learning and Memory, 139, 128-134.
Roth, G., Dicke, U. (2013). Evolution of Nervous Systems and Brains. In: Galizia, C., Lledo, PM. (eds) Neurosciences – From Molecule to Behavior: a university textbook. Springer Spektrum, Berlin, Heidelberg.
Ryoo J, Park S, Kim D. An Inhibitory Medial Preoptic Circuit Mediates Innate Exploration. Front Neurosci. 2021 Aug 23;15:716147.
Santos-Durán GN, Ferreiro-Galve S, Mazan S, Anadón R, Rodríguez-Moldes I, Candal E. Developmental genoarchitectonics as a key tool to interpret the mature anatomy of the chondrichthyan hypothalamus according to the prosomeric model. Front Neuroanat. 2022 Aug 4;16:901451.
Schredelseker T, Driever W. Conserved Genoarchitecture of the Basal Hypothalamus in Zebrafish Embryos. Front Neuroanat. 2020 Feb 6;14:3.
Schwindt P, Crill W. Mechanisms underlying burst and regular spiking evoked by dendritic depolarization in layer 5 cortical pyramidal neurons. J Neurophysiol. 1999 Mar;81(3):1341-54.
Seung HS. How the brain keeps the eyes still. Proc Natl Acad Sci U S A. 1996 Nov 12;93(23):13339-44.
Stephenson-Jones M, Floros O, Robertson B, Grillner S. Evolutionary conservation of the habenular nuclei and their circuitry controlling the dopamine and 5-hydroxytryptophan (5-HT) systems. Proc Natl Acad Sci U S A. 2012 Jan 17;109(3):E164-73.
Stöber, T. M., Lehr, A. B., Hafting, T., Kumar, A., & Fyhn, M. (2020). Selective neuromodulation and mutual inhibition within the CA3–CA2 system can prioritize sequences for replay. Hippocampus, 30(11), 1228-1238.
Suryanarayana SM, Pérez-Fernández J, Robertson B, Grillner S. The Lamprey Forebrain – Evolutionary Implications. Brain Behav Evol. 2022;96(4-6):318-333.
Thirtamara Rajamani K, Barbier M, Lefevre A, Niblo K, Cordero N, Netser S, Grinevich V, Wagner S, Harony-Nicolas H. Oxytocin activity in the paraventricular and supramammillary nuclei of the hypothalamus is essential for social recognition memory in rats. Mol Psychiatry. 2024 Feb;29(2):412-424.
Wee CL, Song EY, Johnson RE, Ailani D, Randlett O, Kim JY, Nikitchenko M, Bahl A, Yang CT, Ahrens MB, Kawakami K, Engert F, Kunes S. A bidirectional network for appetite control in larval zebrafish. Elife. 2019 Oct 18;8:e43775.
Whitlock, J. R., Pfuhl, G., Dagslott, N., Moser, M. B., & Moser, E. I. (2012). Functional split between parietal and entorhinal cortices in the rat. Neuron, 73(4), 789-802.
Wullimann MF. The Neuromeric/Prosomeric Model in Teleost Fish Neurobiology. Brain Behav Evol. 2022;97(6):336-360.
Yamaguchi T, Danjo T, Pastan I, Hikida T, Nakanishi S. Distinct roles of segregated transmission of the septo-habenular pathway in anxiety and fear. Neuron. 2013 May 8;78(3):537-44.
Zylberberg J, Strowbridge BW. Mechanisms of Persistent Activity in Cortical Circuits: Possible Neural Substrates for Working Memory. Annu Rev Neurosci. 2017 Jul 25;40:603-627.




