H.l (lateral hypothalamus) is a key node in the foraging system and has an interesting capability of distinguishing a food zone from a non-food zone [Jennings et al 2015]. In a sense foraging is searching for a food zone and then eating.

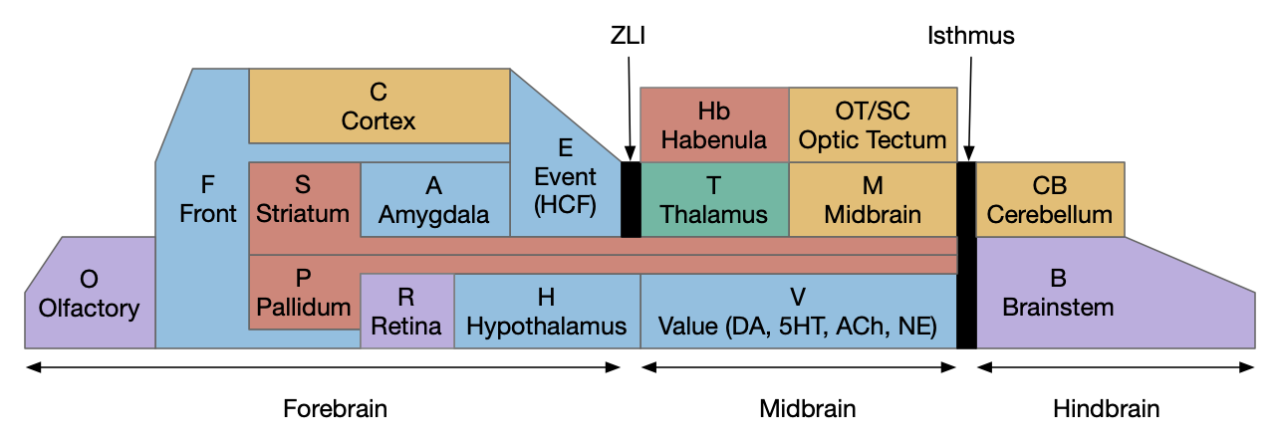
The above diagram shows the foraging phases that I’ve already covered in earlier essays. Importantly, each phase is an independent action path as part of a distributed system, not merely a state in a state machine. To force the separate action paths to act like a state machine, each transition needs to suppress the preceding and following state. In particular the eating phase needs to inhibit the seeking system. This lateral inhibition is important because circuitry is required to force activation of only a single system at a time.
The food zone is particularly interesting for filter feeding, which is naturally area based and long term, as opposed to snapping up a morsel of food. Non-vertebrate chordates are filter feeders, lamprey larvae are filter feeders, and early jawless vertebrates were also likely filter feeders [D’Aniello et al 2023]. Tunicate ascidians, the closest non-vertebrate chordates, have an extreme version of this foraging loop, where the tadpoles find a feeding place after swimming for 12 hours and then settling in place for their adult life [Anselmi et al 2024]. The ascidian foraging state marine is a straight line that ends in the eating phase in the food zone, not continuing in a loop. The ascidian search and settle might give a hint how the vertebrate foraging circuitry is organized.
Ascidians
As covered in essay 30, the ascidian larva nervous system has several seeking (taxis) systems: geotaxis (gravity avoidance – moving up), phototaxis (light avoidance), and dimming for predator and obstacle avoidance. Ascidian navigation disperses the larva from its parent and prefers to settle on the underside of ledges by avoiding gravity while avoiding light. Its settling sensors also avoid toxic or irritating areas and may try to find food-friendly areas, although the specific sensor capabilities aren’t well known. When the larva finds an appropriate place, around 14 hours after hatching, it settles for life [Hoyer et al 2024].

The above diagram is a functional representation of the ascidian larva navigation brain. For this essay the important part is the palp and food-zone sensor and the settling neurons that inhibit motor neurons. The palms are three tentacle-like protrusions from the larva head, which attach the ascidian to a rock with cement glands [Johnson et al 2024]. They contain chemosensory and mechanosensors that distinguish the settling zone from non-settling zones [Hoyer et al 2024]. Interestingly, the genetic markers for the palp neurons are similar to markers for the vertebrate forebrain.
Head cement glands still exist in some fish larvae [Pottin et al 2010] and most frog tadpoles [Nokhbatolfoghahai and Downie 2005], [Rétaux and Pottin 2011], [Sive and Bradley 1996]. Frog tadpoles will swim up and attach to the underside of leaves or to the water surface. This cement gland and settling system may have existed in the pre-vertebrate ancestor and shared for tunicates and vertebrates. Unlike the ascidians the pre-vertebrates likely did not permanently settle. For the sake of this essay, let’s assume they temporarily settled to filter feed in a location and only moved on if filter feeding was unsuccessful or if forced to move by predators, competitors, or environmental hazards.

The above diagram shows specific neurons in the ascidian larva brain. The importance here is the glutamate pnIN (palp interneuron) to GABA pnRN (palp relay neuron), which inhibits all motor neurons and interneurons. Comparing vertebrate and ascidian neural systems is sketchy and probably should be avoided because both have diverged [Holland 2016]. For this essay, I’ll ignore that sound advice to try to motivate part of the vertebrate nervous system.

The above diagram shows the H.stn (subthalamic nucleus) as fulfilling a similar role as the pnIN from ascidian Ciona, suppressing seek in preparation for eating. Part of P.v (ventral pallidum) suppresses S.v (ventral striatum) during eating [Vachez et al 2021]. This P.v “arkypallidal” subset is named after similar neurons in P.ge (globus pallidus) that suppresses S.d (dorsal striatum). Although the driver of this eating suppression isn’t known, the timing of the arkypallidal activation closely matches V.dr serotonin food activation [Spring and Nautiyal 2024], ramping at the end of seek and peaking after eating. Also, H.stn and P.ge form an oscillating pair, evident in Parkinson’s disease. So, it’s plausible that H.stn drives persistent suppression of the seek path in S.v through its projection to P.v, possibly influenced or driven by V.dr (dorsal raphe, serotonin). This specific path is speculation but seems compatible with experiments. The second suppression path is the well-known H.stn to S.nr (substantia nigra pars reticulata) that suppresses motor activity. Snr has a widespread suppression or MLR (midbrain locomotor region), R.rs (reticulospinal motor command), and Snr suppresses Snc (substantia nigra parsa compacta dopamine). Note that the medial H.stn, the area connected with P.v, merges with H.l with minimal boundary [Haynes and Haber 2013].
Food zone
Let’s return to the H.l food zone in [Jennings et al 2015] and consider where the food zone information might come from. Following [Jacobs 2012], let’s treat olfaction as the central sense for navigation, which is particularly compelling for food zones.
The diagram below shows the H.l main connectivity. Not displayed is the H.l internal sensing of nutrient information peptides like glucose sensing and leptin fat sensing. H.l doesn’t receive direct sensory input with the exception of R.pb (parabrachial nucleus), which sends nociceptive information like itch or pain. Because an itchy or painful place is a poor choice for filter feeding, this R.pb input is negative place information for a filter-feeding zone, but R.pb doesn’t give positive reasons to stay like food odors.

As the diagram suggests, the information H.l receives about food sources is very abstract. It receives cue information from A.bl (basolateral amygdala), place information from E.hc (hippocampal complex), value-like information from F.ofc (orbitofrontal cortex) and task-like information from F.vm (ventromedial prefrontal cortex). All of those areas are strongly connected with the olfactory system. While H.l doesn’t receive odor place information directly from sensors, it receives multiple organizational perspectives on odor information. P.bst (bed nucleus of the stria terminalis) receives very similar olfactory input as H.l, and it also receives negative information from R.pb. However, R.pb sends different nociceptive information to the S.a (central amygdala)/P.bst extended amygdala than it sends to H.l [Arthurs et al 2023]. The R.pb projections to H.l compared to S.a/P.bst are not redundant.
Not only are the H.l inputs abstract, but the outputs are also abstract, in contrast to direct action paths. This abstraction might be a later evolutionary development, similar to V.pt (posterior tuberculum) in zebrafish. V.pt is roughly homologous to Vta (ventral tegmental area) in mammals, but V.pt has more direct locomotor output to MLR (midbrain locomotor region), while most of Vta’s output is generally abstract.
As a note, the diagram does not include H.l ox (orexin) or H.l mch (melanin-concentrating hormone), partially for simplicity and partially because the zebrafish H.l is distinct from the ox and mch populations, suggesting that the mammalian ox and mch areas of H.l can be separated from the rest of H.l function. The diagram also omits some other connections like Ppt (pedunculopontine nucleus).
Food and serotonin
Returning to the foraging state diagram, it’s important that each “state” is a large, distributed, complex system, not a state in a state machine. The seek state includes areas like S.v, Vta, H.l, E.hc, F.pfc, and the motor regions MLR and R.rs (reticulospinal motor command) with the help of cortical areas and can include OT (optic tectum). Although the eating state is small, it is still comprised of many areas, including V.dr (dorsal raphe), OT.d, R.my.irt (medulla eating), H.l, H.pstn (parasubthalamic nucleus), R.pb and possibly some Vta and S.v subareas. Although the system is not a state machine, each “state” needs to laterally suppress the other systems to prevent multiple action paths from colliding.

The split between eat and seek is important, because many studies merge the behavior into a general category “feeding.” Because some experiments only measure total feeding, it can be difficult to distinguish whether the experiment is measuring a seek effect or an eating effect. For example, eating needs to suppress seek to keep the animal from wandering away from the food. If an experiment stimulates eat but inhibits seek, the animal might not search for food even if it’s ready to eat. If it doesn’t seek food, it doesn’t find food.
This distinction between eating and seeking is exhibited by the question of serotonin, which is a heterogeneous system that has a role in feeding. The serotonin from V.dr is a heterogenous system with V.dr having at least 14 different genetic clusters [Okaty et al 2020] with at least 11 different projection patterns [Ren et al 2014]. Earlier studies noted that 30% of V.dr were active during eating [Fornal et al 1996], and many others have noted V.dr being active for “reward” (eating).
Suppose one component of V.dr serotonin encourages eating while discouraging seeking. If an experiment floods the brain with serotonin, it might see total feeding drop because serotonin suppresses seeking food, even if it encourages long meals when it finds food. The confusion becomes greater for studies looking for the even more abstract “reward” as opposed to concrete eating. The point being that serotonin in particular is a complicated system, not reducible to a single value or function.

The above diagram shows some of the eating-related projections. Only a few of the 14 V.dr subtypes are know. The V.dr to Vta connection is one of the known projections and drives the seek system [Courtiol et al 2021], [Wang HL et al 2019]. Unfortunately, the other projections are not known, in particular the 30% of V.dr that is active while eating [Bromberg-Martin et al 2010].
V.dr enhances satiety with 5HT2c.q (serotonin G-q stimulating receptor) in H.arc POME satiety neurons, which suppresses the AgRP hunger peptide. Note that AgRP drops just before eating, suggesting that it’s a seek-promoting system, but an eating-promoting system [Bhave and Nettow 2021]. The prediction suppression only occurs after training and V.dr serotonin shows inverse behavior, possibly suggesting V.dr as suppressing H.arc. Untrained V.dr serotonin only responds after tasting [Li et al 2016], but trained V.dr serotonin responds about 2 seconds before eating [Zhong et al 2016].
Filter feeding and foraging theory
Let’s the consider filter feeding using foraging theory. Foraging theory studies how animals browse patches of food, such as a cluster of flowers for a bee or worms in pine cones for birds [Krebs et al 1974] or a hunting spot for a predator. In particular, foraging theory considers how long the animal should stay at a particular patch before deciding to move on: measuring the give up time. A filter-feeding proto-vertebrate needs to decide if the current food rate is good enough to stay at the current food zone.
The MVT (marginal value theorem) suggests that an animal should move on if the current patch has less food than the environment average [Charnov 1976]. MVT has simplifying assumptions that are challenged by the complexity in the world [Pyke 1984], [Wajnberg et al 2006]. MVT assumptions include omniscience, immortality, determinism, no competition, no predation, and no hunger. Some of those complexities are important to the essay, particularly the omniscience. In MVT the animal knows the average environment food value, but this omniscience isn’t plausible for simple animals [Tenhumberg et al 2001], and the essay animal has almost no learning at all. Realistic search is stochastic and can fail, such as a predator hunting, which is particularly important if the animal is starving. Starvation and satiation are also not covered by the MVT. If the animal is starving, it might stick with a non-optimal, low quality food source below the environment average because not finding a better patch is too risky. Simple organisms use rules of thumb instead of complex strategy, and even birds seem to use a constant give up time [Krebs et al 1974].
As a side note, the foraging terms for eating (“exploiting”) and searching for a new patch (“exploring”) have been appropriated by RL (reinforcement learning) [Sutton and Barto 2018] with some differences in meaning. Reinforcement learning use an n-armed bandit (gambling slot machine) model, where exploring means finding the reward rates of the other arms before deciding on the best arm to exploit. The RL focus is on gather information, generally in a finite and persistent system. In contrast, this essay uses the original foraging terminology.
Covered in essay 36, vertebrate food motivation divides into hunger-driven (“homeostatic”) and opportunistic (“hedonic”) foraging. These form two levels of search and involve different circuits with some overlap. When no longer hungry, mice will not eat plain food but will still eat rich food. In terms of foraging theory, hungry mice will stay longer at poor patches, while sated mice will leave more quickly.
Simulation complexity
After starting to implement the simulation, the issue of complication became overwhelming. Specifically, adding the striatum is too complicated. Consider the issue of distinguishing the eating function of dopamine vs serotonin, when both are responsive to eating food. That similarity makes it difficult to find the system function. The system must have developed from a simpler system because the ascidian feeding or amphioxus feeding is not overly complicated. For the sake of the simulation, I’m backing off and considering only the hindbrain and hypothalamus systems, treating the striatum as a later enhancement.
Hypothalamus and raphe nuclei
The core of the simulation is the pair of H.l and V.dr. As mentioned above, H.l is driven by food zone indicators and can drive both seeking and eating. V.dr is responsive to eating and as part of the hindbrain (it derives from r1) it is a good candidate for primitive, tunicate-like filter feeding circuitry.

The diagram above is a simplification, where the Ob to H.l connection represents an ancient version of the food zone system. The V.dr to R.nts (nucleus of the solitary tract) connection includes more hindbrain structures such as medulla eating circuits. The simplification has H.l as a food zone controller and V.dr as an eating sustaining manager.
Although V.dr is a serotonin system, not V.dr neurons are non-serotonin, both glutamate and GABA. As mentioned above the V.dr and V.mr (median raphe) serotonin neurons have at least 11-14 distinct neuron types and projection types. For the essay I’m assuming at least one serotonin neuron type is a measure of eating food. In the simulation successful filter feeding increases the serotonin for eating.
Start and sustain
Let’s return to foraging, where the central decision is when to stop exploiting a patch if it’s not effective. Consider a simple where the animal gives up on a patch if the feeding rate drops below a fixed threshold. Filter feeding naturally has delays between starting filter feeding, trapping some prey, and later receiving nutrients in the gut. This raises a problem: the feeding rate is zero until some food is digested, which implies the animal should give up immediately.

One solution is to prime the system with a start signal. While the start signal exists, the animal won’t leave even if it hasn’t digested any nutrients. In the simulation H.l is responsible for the start signal and V.dr is responsible for both the sustain and for integrating the two systems. The H.l start signal comes from the food zone detection.
However, the start signal raises a new issue because the start signal must stop to allow sustain to act as the primary decision variable. If H.l always sends the food zone signal to V.dr, it will remain active as long as the animal is in the food zone, preventing the animal from leaving the zone. So, H.l itself needs a timeout. The simulation uses a striatum timeout to disable the H.l food zone signal. The striatum connection can either represent the striatum layer between the olfactory and cortical layers and H.l, or it can represent H.l reciprocal input to the striatum.
The start timeout has the same issues as other striatum systems. Specifically, it needs to remain timed out until the animal leaves the food zone.
Simulation
The screenshot below shows the animal feeding from a low-quality food zone. The grey star is a food zone (grey represents poor food). The nearby purple checkerboard is an avoidance zone, representing an aversive area such as itch or high carbon dioxide.

In the screenshot the startup signal from H.l is temporarily sustaining feeding. It will soon timeout and the animal will abandon the food zone.
Avoidance response and search
The simulation adds two other serotonin-based systems: one for avoiding toxic areas and one for search. Avoidance is one of the V.mr functions. The search serotonin represents the V.dr to Vta connection, despite the current essay disabling the seek function. These two functions may not be serotonin functions because V.mr avoidance is largely non-serotonin, and the V.dr to Vta connection is primarily glutamate. Because the avoidance and search are not the primary focus of the essay, I’m putting off the question of accuracy to a later essay.
Discussion
The essay’s big questionable decision is the omission of the striatum, particularly because I’ve already used the striatum for give-up timing. For eating as opposed to seeking, one possible area appears to be S.dl.vl, which is the orobranchial, mouth area [Foster et al 2021]. Because S.dl receives late dopamine from food in the gut, it might be a good candidate for filter feeding sustain.

A second area is S.msh.d (dorsal medial shell) which responds to hedonic “liking” and drives strong eating [Castro et al 2016], [Richard and Berridge 2011], [Richard et al 2013]. S.msh.d drives H.l, which is central to the essay. In addition S.msh has longer, sustained dopamine (5-10s) contrasted with shorter dopamine in S.dl (100ms) [de Jong et al 2022].
From a motivational perspective, S.dl.vm and S.msh.d are strong candidates, but they lack the lateral inhibition of seek that’s necessary for the state machine to work. S.dl.vl also works through OT.d.l (optic tectum deep motor areas), which would add more complexity to this essay. In contrast the V.dr serotonin is already part of the hindbrain motor areas, and serotonin is already inhibitory toward seek. V.dr requires fewer additional systems to work. For future work, the two striatum areas are strong areas to research.
References
Anselmi C, Fuller GK, Stolfi A, Groves AK, Manni L. Sensory cells in tunicates: insights into mechanoreceptor evolution. Front Cell Dev Biol. 2024 Mar 14;12:1359207.
Arthurs JW, Pauli JL, Palmiter RD. Activation of Parabrachial Tachykinin 1 Neurons Counteracts Some Behaviors Mediated by Parabrachial Calcitonin Gene-related Peptide Neurons. Neuroscience. 2023 May 1;517:105-116.
Bhave VM, Nectow AR. The dorsal raphe nucleus in the control of energy balance. Trends Neurosci. 2021 Dec;44(12):946-960.
Bromberg-Martin ES, Hikosaka O, Nakamura K. Coding of task reward value in the dorsal raphe nucleus. J Neurosci. 2010 May 5;30(18):6262-72.
Castro DC, Cole SL, Berridge KC. Lateral hypothalamus, nucleus accumbens, and ventral pallidum roles in eating and hunger: interactions between homeostatic and reward circuitry. Front Syst Neurosci. 2015 Jun 15;9:90.
Charnov, E. L. (1976b). Optimal foraging: The marginal value theorem. Theoretical Popula- tion Biology, 9, 129–136.
Courtiol E, Menezes EC, Teixeira CM. Serotonergic regulation of the dopaminergic system: Implications for reward-related functions. Neurosci Biobehav Rev. 2021 Sep;128:282-293.
D’Aniello S, Bertrand S, Escriva H. Amphioxus as a model to study the evolution of development in chordates. Elife. 2023 Sep 18;12:e87028.
de Jong JW, Fraser KM, Lammel S. Mesoaccumbal Dopamine Heterogeneity: What Do Dopamine Firing and Release Have to Do with It? Annu Rev Neurosci. 2022 Jul 8;45:109-129.
Fornal CA, Metzler CW, Marrosu F, Ribiero-do-Valle LE, Jacobs BL. A subgroup of dorsal raphe serotonergic neurons in the cat is strongly activated during oral-buccal movements. Brain Res. 1996 Apr 15;716(1-2):123-33.
Foster NN, Barry J, Korobkova L, Garcia L, Gao L, Becerra M, Sherafat Y, Peng B, Li X, Choi JH, Gou L, Zingg B, Azam S, Lo D, Khanjani N, Zhang B, Stanis J, Bowman I, Cotter K, Cao C, Yamashita S, Tugangui A, Li A, Jiang T, Jia X, Feng Z, Aquino S, Mun HS, Zhu M, Santarelli A, Benavidez NL, Song M, Dan G, Fayzullina M, Ustrell S, Boesen T, Johnson DL, Xu H, Bienkowski MS, Yang XW, Gong H, Levine MS, Wickersham I, Luo Q, Hahn JD, Lim BK, Zhang LI, Cepeda C, Hintiryan H, Dong HW. The mouse cortico-basal ganglia-thalamic network. Nature. 2021 Oct;598(7879):188-194.
Haynes WI, Haber SN. The organization of prefrontal-subthalamic inputs in primates provides an anatomical substrate for both functional specificity and integration: implications for Basal Ganglia models and deep brain stimulation. J Neurosci. 2013 Mar 13;33(11):4804-14.
Holland, L. Z. (2016). Tunicates. Current Biology, 26(4), R146-R152.
Hoyer J, Kolar K, Athira A, van den Burgh M, Dondorp D, Liang Z, Chatzigeorgiou M. Polymodal sensory perception drives settlement and metamorphosis of Ciona larvae. Curr Biol. 2024 Mar 25;34(6):1168-1182.e7.
Jacobs L. F. (2012). From chemotaxis to the cognitive map: the function of olfaction. Proc. Natl. Acad. Sci. U.S.A. 109(Suppl. 1) 10693–10700 10.1073/pnas.1201880109
Jennings JH, Ung RL, Resendez SL, Stamatakis AM, Taylor JG, Huang J, Veleta K, Kantak PA, Aita M, Shilling-Scrivo K, Ramakrishnan C, Deisseroth K, Otte S, Stuber GD. Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell. 2015 Jan 29;160(3):516-27.
Johnson CJ, Razy-Krajka F, Zeng F, Piekarz KM, Biliya S, Rothbächer U, Stolfi A. Specification of distinct cell types in a sensory-adhesive organ important for metamorphosis in tunicate larvae. PLoS Biol. 2024 Mar 13;22(3):e3002555.
Krebs JR, Kacelnik TP (1978) Tests of optimal sampling by foraging great tits. Nature 275:27–31
Li Y, Zhong W, Wang D, Feng Q, Liu Z, Zhou J, Jia C, Hu F, Zeng J, Guo Q, Fu L, Luo M. Serotonin neurons in the dorsal raphe nucleus encode reward signals. Nat Commun. 2016 Jan 28;7:10503.
Nokhbatolfoghahai M, Downie JR. Larval cement gland of frogs: comparative development and morphology. J Morphol. 2005 Mar;263(3):270-83. doi: 10.1002/jmor.10305.
Okaty BW, Sturrock N, Escobedo Lozoya Y, Chang Y, Senft RA, Lyon KA, Alekseyenko OV, Dymecki SM. A single-cell transcriptomic and anatomic atlas of mouse dorsal raphe Pet1 neurons. Elife. 2020 Jun 22;9:e55523.
Pottin K, Hyacinthe C, Rétaux S. Conservation, development, and function of a cement gland-like structure in the fish Astyanax mexicanus. Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17256-61.
Pyke, G.H., 1984. Optimal foraging theory: a critical review. Annual review of ecology and systematics, 15, pp.523-575.
Ren J, Isakova A, Friedmann D, Zeng J, Grutzner SM, Pun A, Zhao GQ, Kolluru SS, Wang R, Lin R, Li P, Li A, Raymond JL, Luo Q, Luo M, Quake SR, Luo L. Single-cell transcriptomes and whole-brain projections of serotonin neurons in the mouse dorsal and median raphe nuclei. Elife. 2019 Oct 24;8:e49424.
Rétaux S, Pottin K. A question of homology for chordate adhesive organs. Commun Integr Biol. 2011 Jan;4(1):75-7.
Richard JM, Plawecki AM, Berridge KC. Nucleus accumbens GABAergic inhibition generates intense eating and fear that resists environmental retuning and needs no local dopamine. Eur J Neurosci. 2013 Jun;37(11):1789-802.
Richard JM, Berridge KC. Nucleus accumbens dopamine/glutamate interaction switches modes to generate desire versus dread: D(1) alone for appetitive eating but D(1) and D(2) together for fear. J Neurosci. 2011 Sep 7;31(36):12866-79.
Sive H, Bradley L. A sticky problem: the Xenopus cement gland as a paradigm for anteroposterior patterning. Dev Dyn. 1996 Mar;205(3):265-80.
Spring MG, Nautiyal KM. Striatal Serotonin Release Signals Reward Value. J Neurosci. 2024 Oct 9;44(41):e0602242024.
Sutton, R. S., & Barto, A. G. (2018). Reinforcement learning: An introduction (2nd ed.). The MIT Press.
Tenhumberg, B., Keller, M. A. & Possingham, H. P. Using Cox’ s proportional hazard models to implement optimal strategies: an example from behavioural ecology 2. Wasp behaviour model. Behaviour 33, 597–607 (2001).
Vachez YM, Tooley JR, Abiraman K, Matikainen-Ankney B, Casey E, Earnest T, Ramos LM, Silberberg H, Godynyuk E, Uddin O, Marconi L, Le Pichon CE, Creed MC. Ventral arkypallidal neurons inhibit accumbal firing to promote reward consumption. Nat Neurosci. 2021 Mar;24(3):379-390.
Wajnberg, E., Bernhard, P., Hamelin, F. & Boivin, G. Optimal patch time allocation for time-limited foragers. Behav. Ecol. Sociobiol. 60, 1–10 (2006).
Wang HL, Zhang S, Qi J, Wang H, Cachope R, Mejias-Aponte CA, Gomez JA, Mateo-Semidey GE, Beaudoin GMJ, Paladini CA, Cheer JF, Morales M. Dorsal Raphe Dual Serotonin-Glutamate Neurons Drive Reward by Establishing Excitatory Synapses on VTA Mesoaccumbens Dopamine Neurons. Cell Rep. 2019 Jan 29;26(5):1128-1142.e7.
Zhong W, Li Y, Feng Q, Luo M. Learning and Stress Shape the Reward Response Patterns of Serotonin Neurons. J Neurosci. 2017 Sep 13;37(37):8863-8875.


























