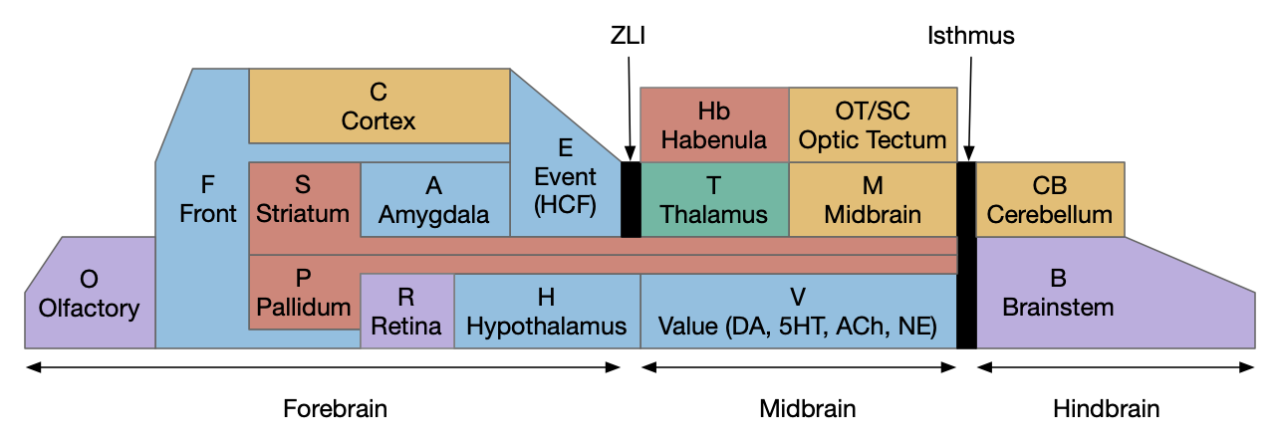
Early studies described the hindbrain as a “reticular formation,” which means that under the microscope it looks like a find netlike structure, as if it was an undifferentiated, impenetrable mass. However, using modern genetic tools, the hindbrain shows a strongly hierarchical organization [Vishwanathan et al 2024]. The hindbrain is capable of action decision-making on its own, in opposition to a theory that forebrain systems like the basal ganglia are primary for action decision [Humphries et al 2007].
The vertebrate hindbrain can also be explored as a sequence of evolutionary additions, where early hindbrain provides locomotion but not other functions, as in the non-vertebrate chordate Amphioxus. The subcircuits of the hindbrain could be describes as a sequence:
- Locomotion – exists in Amphioxus
- Filer feeding – added in tunicates like the ascidian Ciona
- Vestibular corrections for locomotion (“posture”) – lamprey larva
- Vestibular stabilization for image vision – lamprey adult
In this essay I’m reviewing the hindbrain as this sequence of evolutionary steps and using it to introduce decision-making circuit patterns and also to clarify the hindbrain capabilities, which allow higher areas to use its fundamental abilities.
Xenopus tadpole
As a study animal the Xenopus (African clawed frog) tadpole has advantages of a simple nervous system and the additional advantage of similar behaviors to the chordate ascidian tadpole, believed to be the closest chordates to the vertebrates. The ascidian tadpole swims for only a few hours before permanently attaching to a rock or overhang using a cement gland on its palps (basically head) and settling permanently as a sessile filter feeder. It uses a simple photoreceptor ocellus for phototaxis and a gravity-sensing otolith for geotaxis to first swim up, then down if necessary, but preferably swimming up to a shadow-producing ledge. The Xenopus tadpole in an early stage also swims upward beneath a shadow to attach its cement gland to the underside of a leaf [Jamieson and Roberts 2000], [Roberts et al 2000], [Nokhbatolfoghahai and Downie 2005], [Rétaux and Pottin 2011] and some amphibian species tadpoles filter feed [Pryor 2014], but at a later development stage.
Amphibian locomotor CPG
The swimming CPG (central pattern generator) produces the alternating left and right muscle movements that progresses toward the tail along the trunk segments. The caudal hindbrain (R5-R8) can sustain swimming oscillations, even if more rostral areas are lesioned [Soffe et al 2009]. In contrast to the rhythmic caudal hindbrain, the rostral hindbrain has only loose rhythm and not synchronized to swimming [Soffe et al 2009]. So, along with locomotion, this circuit can demonstrate persistent neural activity as in working memory and show that persistence does not necessarily imply a single attractor state.
The caudal hindbrain (R5-R7, aka medulla) and N.sp (spinal cord) locomotor circuits have a simple organization with four neuron types [Soffe et al 2009].
- N.mn (motor neuron) drives the muscle
- R.din (descending interneuron) pattern generator and premotor
- R.cin (commissural interneuron) inhibits opposite side to organize anti-phase oscillation
- R.ain (ascending interneuron) lateral inhibition of sensory input [Roberts et al 2010]

R.din is the primary premotor neuron and its cellular properties support oscillations both intrinsically and with rebound firing after inhibition. R.din are the most medial and ventral stripe in the hindbrain and are marked by genetic transcription factor chx10 [Li WC and Soffe 2019]. The chx10 locomotor neurons are strongly conserved in vertebrates. In mammals chx10 hindbrain neurons contribute to locomotion, stopping [Bouvier et al 2015] and turning [Huang et al 2013], [Cregg et al 2020].
R.din fire rhythmically for swimming and fire early in the swimming cycle [Soffe et al 2009]. R.din initiate swimming and drive the rest of the CPG [Ferrario et al 2021]. R.din typically fire a single AP (action potential) on each swimming cycle alternative left and right [Li WC and Soffe 2019]. The early tadpole R.din population size is approximately 50 neurons [Soffe et al 2009].
R.din can oscillate through a combination of:
- Intrinsic NMDA-dependent oscillation [Li WC et al 2010], [Moult and Cottrell 2013], [Bubnys et al 2019]
- Rebound firing after R.cin inhibition [Soffe et al 2009], [Li WC et al 2010], [Moult and Cottrell 2013], [Svensson et al 2017], [Koutsikou et al 2018]
- R.din electrical coupling [Li WC et al 2010]
The R.din intrinsic NMDA-dependent oscillation can produce membrane oscillations even when APs are blocked [Li WC et al 2010], likely dependent on sodium leak currents [Svensson et al 2017]. Leak currents modulate AHP (afterhyperpolarization) and ADP (afterdepolarization), which regulates the membrane potential after an AP, which either makes subsequent APs harder to fire or easier to fire. So, cell properties can either encourage persistent firing or force only a single AP spike with a long refractory period. For R.din these properties promote oscillation, which differs from the other neurons in this circuit, which do not promote oscillations.
R.din intrinsic oscillation appears to be a secondary effect, not the main CPG driver [Bubnys et al 2019]. Rebound firing after R.cin inhibition is a stronger effect [Soffe et al 2009], [Moult and Cottrell 2013]. Although R.din oscillate independently, they will typically fire a single AP on each swimming cycle, alternating left and right [Li WC and Soffe 2019]. In the lamprey, there is a debate whether the commissural couple of the CPG is necessary for oscillation or if each side is an independent oscillator [Cangiano and Grillner 2018], [McClellan 2018]. Stimulating single R.din neurons in tadpoles rarely initiates swimming, with exceptions when the hindbrain is disconnected from the midbrain [Li WC and Soffe 2019], suggesting that the population of R.din is essential.
R.ci inhibits the contralateral R.din to coordinate the paired oscillators to fire in opposite phases. In Xenopus R.cin are glycine interneurons. As described above, the R.cin inhibition of R.din drives the oscillator by promoting rebound APs.
R.ain are inhibitory interneurons that suppress sensory input [Roberts et al 2010]. These form lateral inhibition with other CPGs, blocking sensory input to discourage competing actions. While the tadpole is swimming, it will respond less to touching of the trunk and tail.
While the CPGs for swimming are relatively simple, coordinating the multiple limbs for tetrapods is more complicated, particularly with the added challenge of gravity. For mammals and likely all tetrapods, the rhythmic generators are correspondingly more complicated. The exact coupling of rhythm generators and multiple pattern generators is a debated [McCrea and Rybak 2008].
A few things to take away here. First, the feedback inhibition in this CPG produces oscillation, which will be important for later WTA (winner-take-all) decision-making, because decision-making uses a similar structure. Second, the oscillatory sustaining behavior is dependent on intrinsic cell properties, not simply circuit connectivity. The R.din intrinsic oscillation and inhibition rebound are specific properties that neighboring neurons lack. They are specifically designed for their roles in driving oscillation.
Swimming eigenshapes
Borrowing from linear algebra, movement for simple bilateral animals can be described by a small number of basic vectors that encode basic shapes “eigenshapes,” implying that swimming actions are low dimensional. Almost all (95%) locomotion for the flatworm C. elegans can be described by four dimensions [Stephens et al 2008], almost all (96%) of ascidian tadpole swimming can be described by six vectors [Athira et al 2022] and 96% of zebrafish larva swimming is captured by three vectors [Girdhar et al 2015]. These basis vectors encode the lateral displacement of each body segment from the midline and a linear combination of the vectors produces the majority of posture shapes while swimming.
The zebrafish larva has fewer dimensions because it has a stiff spine that limits posture variability [Girdhar et al 2015]. In zebrafish larva, two of the three dimensions encode the swimming oscillation and the third encodes an initial turn. After an initial turn, zebrafish settle into a stereotypical swimming pattern. Most of the variation in swimming bouts is attributable to the depth of the initial of the initial turn.
The low dimensionality of vertebrate swimming suggests that it uses relatively simple circuits. The swimming CPG already covers two of the three basis shapes with variation in the initial turn using distinct action paths. This division between a stereotyped forward movement modulated by distinct turning circuits persists in tetrapods. The MLR (midbrain locomotor region) produces forward motion whether it’s stimulated unilaterally or bilaterally in mammals [Brocard et al 2010] and salamanders [Ryczko et al 2016], while separate turning systems drive chx10 R.rs (reticulospinal) neurons in the central hindbrain [Cregg et al 2020].
Tadpole locomotion action paths
At hatching tadpoles have limited senses, which simplifies understanding their action paths. Tadpoles have touch sensitivity, light sensitivity from the pineal eye, and possibly water movement from the lateral line [Roberts et al 2010]. The lateral eyes are not functional until a few days later.
Tadpoles have four major action paths [Borisyuk et al 2017];
- Dimming from pineal eye, producing upward swimming [Jamieson and Roberts 2000], [Larbi et al 2022]
- Head press, which stops swimming when the tadpole touches a ceiling [Roberts et al 2010]
- Head touch, which drives swimming [Buhl et al 2015], [Koutsikou et al 2018]
- Trunk touch, which drives swimming [Roberts et al 2014], [Koutsikou et al 2018]
Beside the four main action paths, tadpoles have a few additional action paths:
- Struggling from prolonged touch on both sides [Soffe 1991], [Roberts et al 2010]
- UV avoidance from a deep brain UV photoreceptor, independent of retina and pineal eye [Currie et al 2016]
- Stopping swimming using ATP as a timeout [Dale 1998]

The above diagram shows the action paths and their implementing circuits. Each box represents a small collection of functionally-similar neurons. As a general pattern, primary sensory neurons like N5 (trigeminal head touch, 5th cranial nerve) drive a collection of relay neurons like R.in5 (trigeminal interneurons), which spatially integrate the sensory evidence and drive decision pre-motor areas like the CPG R.din neurons or feed into an evidence integrator like the proposed R.xin (extended hindbrain interneurons).
Head press stop
The tadpole has different reactions to head press than for head touch. While head touch drives swimming, head press stops the tadpole when it reaches an attachment site, like the chordate ascidian tadpole [Johnson et al 2024]. Xenopus tadpoles have a cement gland on their head [Nokhbatolfoghahai et al 2005], which they use to attach to the bottom of ledges, lily pads, or the water surface. Similarly the ascidian sensory neurons in the palp area trigger stopping and settling, when the ascidian settles down to its adult sessile filter feeding form. However, the ascidian palp are chemosensory as well as mechanosensory [Ryan et al 2016], [Hoyer et al 2024] and have forebrain transcription factors [Cao et al 2019], [Liu and Satou 2019], but the Xenopus tadpole is purely mechanosensory using hindbrain N5 (trigeminal) [Roberts et al 2010] and no forebrain olfactory input. In addition, the vertebrate N5 trigeminal placode does not exist in tunicates. These differences make it unclear if the prototypical vertebrate was more like the ascidian tadpole or the amphibian tadpole.
Head press neurons are N5 that contact R6.mhr (mid-hindbrain GABA inhibitory) neurons that contact R6.din swimming CPG [Roberts et al 2010]. R6.mhr fire at very low rates without stimulation. R6.mhr cell bodies that respond to head press are in approximately R6-R8 with short dorsal dendrites and long ventral dendrites [Perrins et al 2002]. A single R6.mhr can halt swimming, but single neuron firing is rare because a head press activates all R6.mhr [Ferrario et al 2021]. R6.mhr are rhythmically inhibited by swimming [Perrins et al 2002].
The Xenopus tadpole is non-feeding at this stage, and it loses the head press shortly afterward. Although some amphibian tadpoles do have a filter feeding stage, I haven’t read of any that use the cement gland as part of their filter feeding. The missing chemo sensation and associated forebrain input is not necessary if the animal doesn’t use that information for filter feeding.
Head touch
The tadpole head touch is a distinct action path from the head press. Head touch produces swimming in a random direction. A head touch activates 70 N5 neurons that active R2.in5 (trigeminal interneurons) in R2 and R3, which then activate proposed integrating neurons R.xin [Buhl et al 2012], [Buhl et al 2015]. The R2.in5 are not specific to a head location but integrate from most touch neurons across the ipsilateral head. When the signal is strong enough, nearly all R2.in5 spike at least once. Each R2.in5 connection to R6.din is weak, with a small EPSP and unreliable 49% probability of glutamate. However, because the population does strongly converge to R5.din, simultaneous population firing will drive R5.din and swimming, but a single noisy R5.in5 will not trigger a false alarm.
Even a reflex action requires some nuance. Because individual neurons are noisy and the world itself is noisy, a small or noisy sensation shouldn’t trigger an expensive escape. In the tadpole, N5 head touch strongly converges on R2.in5 simultaneously [Buhl et al 2012].
The swimming direction from a head touch is more complicated than a reflex [Buhl et al 2015]. A strong touch will drive fast 25ms ipsilateral swimming that is reflex-like with a higher threshold and immediate decay, but weaker touches will drive a longer bilateral integration that ramps from 25ms to 50ms, possibly using the proposed R.xin neurons [Koutsikou et al 2018], [Ferrario et al 2021].
Trunk touch
Tadpole trunk touch also drives swimming and is functionally similar to head touch, but is a distinct action path. Head touch neurons are from the trigeminal placode through the N5 cranial nerve, while trunk touch are non-placode Rohon-Beard touch cells that travel through the spine. The trunk sensory pathway goes through N.sp.dlc (dorsolateral commissural) and N.sp.dla (dorsolateral ascending) neurons and then to R.xin integrating neurons, which activates R.din swimming CPG [Borisyuk et al 2017]. N.sp.dlc produces weak, long NMDA excitation of all contralateral neuron types, including motoneurons [Roberts et al 2010]. The swimming direction is unpredictable even if the midbrain is disconnected by a MHB lesion [Ferrario et al 2021].
Although N.sp.dla and N.sp.dlc trunk touch neurons have fast responses that decay rapidly, the R.din CPG neurons show a slow ramping EPSC, which is too long to come only from the trunk sensory neurons [Koutsikou et al 2018]. N.sp.dlc does not appear to contact R.din directly, and N.sp.dlc has a short response on the order of 60ms to 160ms, but R.din shows ramping EPSC for nearly 1s before reaching firing threshold. A hypothetical population of R.xin neurons are proposed to produce the sustained and ramping input. The slower excitation allows time for temporal summation and integration with other inputs such as the pineal eye or head touch, and enables random swimming direction, which improves escape from predators by reducing predictability. These neurons have not yet been identified, but they appear to be hindbrain-specific because a MHB lesion does not eliminate the ramping behavior.
This kind of ramp-to-threshold decision making is commonly modeled as a DDM (drift diffusion model) and studied for decision making in the cortex and OT (optic tectum). The R.xin neurons, when they are identified, could provide insights into cortical decision making by providing a simpler but complete hindbrain circuit in contrast to the more complex, distributed OT and cortex model, which includes complex loops with the thalamus and basal ganglia.
Dimming response
The early Xenopus tadpole (stage 37) doesn’t have retina vision until lateral development (stage 44) after a few days. Until the paired eyes are available, light response uses the single pineal eye, which is closely related to the adjacent habenula. Dimming from M.pineal (pineal eye) causes upward swimming [Jamieson and Roberts 2000]. Upward swimming is in a spiral, like ascidian swimming. The Xenopus tadpole’s upward swimming is like ascidian tadpole upward swimming response to dimming [Bostwick et al 2020], which uses a combination of geotaxis from an otolith and phototaxis from photoreceptors [Olivo et al 2021]. M.pineal drives sensory relay M.dmd (diencephalic/mesencephalic descending neurons), which drives R6.din [Borisyuk et al 2017].
The probability of tadpole responding to light dimming is low. Swimming and reattachment of the cement gland can be quick (8s) but also longer (61s). At stage 44 tadpoles swim continuously, but the pineal eye persists beyond stage 44. At stage 45 lateral eyes become functional. Because young tadpoles do not feed at this stage [Jamieson and Roberts 2000], this swimming and cement-gland attachment is defensive, not for filter feeding.
The Xenopus tadpole also has UV light avoidance using deep UV photoreceptors near the hypothalamus. These photoreceptors are not related to either the retina or to the pineal eye. The action path uses neurons in the caudal hindbrain (R6 area) [Currie et al 2016].
Struggle
Struggle is a distinction motion from swimming, used to escape predator grasp. In Xenopus the structure circuit has a similar structure to a swimming CPG but is a distinct neuron population [Borisyuk et al 2017], [Roberts et al 2010]. While the struggle network is active, the R.din swimming network is inactive.
ATP-based stopping
The previous actions paths covered swimming, but a control system is also necessary for stopping. ATP and adenosine are natural for timing because neural activity uses ATP, which degrades into adenosine. As activity continues adenosine builds up. An adenosine receptor can then inhibit swimming after adenosine builds up. In Xenopus stopping of swimming uses this adenosine/ATP timing system [Dale 1998], [Dale 2002]. When adenosine receptors are disabled, swimming extends from 215s to 600s.
Zebrafish locomotion
The zebrafish larva has additional hindbrain locomotion systems that illustrate hindbrain decision making. The zebrafish hindbrain locomotion sorts into three major clusters: forward translation, left turns, and right turns [Feierstein et al 2023]. Turning uses a common set of chx10 neurons in R.rs.v (ventral reticulospinal) for phototaxis, OMR (optomotor reflex), dark-flash, and spontaneous turns [Huang et al 2013]. Fast escape and slow swimming use distinct circuits, which parallels the fast and slow swimming circuits of the chordate Amphioxus [Lacalli and Candiani 2017]. Neurons in the fast path are more ventral and are born earlier than slower swimming circuits [Agha et al 2024].
The zebrafish escape has at least two independent circuits [Marquart et al 2019]
- R4.mc (Mauthner cell) escape SLC (short-latency C-start)
- R1.llc (long latency C-start)
The R4.mc are giant, easily identified neurons in R4, which are part of the ASR (acoustic startle response), where N8 (auditory nerve) directly drives R4.mc, which directly drive N.sp.mn (spinal motoneurons) in a fast, three synapse escape reflex to loud sounds. Although R4.mc is most studied for its acoustic response, it also drives escape for looming threats from OT [Martorell and Medan 2022], and touch from the trunk and head [Bhattacharyya et al 2017]. Early in development, head touch neurons are the first R4.mc input [Kohashi et al 2012].
R1.llc are a set of R1 neurons that respond to slower environmental threats and also produce escape swimming [Marquart et al 2019]. While R4.mc are extremely well-studied, the connectivity and function of R1.llc is less known beyond their response to lower priority threats. The threshold between R4.mc SLC and slower R1.llc LLC is modulated by many factors, including specific calcium ion receptors in R6.dc [Shoenhard et al 2022].
The ascidian tadpole has ddN cells, which are proposed as homologous to R4.mc. ddN cells are the only descending and decussating (commissural) cells in Ciona [Ryan et al 2017].
In zebrafish larva locomotion also initiates from M.nmlf (nucleus of the medial lateral fasciculus). The mlf is the main locomotion and oculomotor nerve tract through the hindbrain. M.nmlf is closely tied to M.pt (pretectum) and OT (optic tectum) and drives movement for optic flow, dimming, and hunting. M.nmlf is also activated by head touch [Sankrithi et al 2010], serving a similar function to the Xenopus R.in5 neurons.
Mauthner cell – decision with FFI
The R4.mc (Mauthner cell) escape circuit is a good illustration of decision making, because the circuit rapidly chooses between turning left or right. R4.mc escape circuit is only three synapses long, with the N8 (acoustic) sensory neurons, R4.mc premotor neurons, and the final N.sp.mn (spinal) motoneurons. R4.mc choose an initial turn direction, suggesting that only one of the two R4.mc cells should activate to avoid delays from competing, simultaneous left and right activation. Although touch may be a simple, non-conflicting system, where touching the left of the head drives a right turn, the direction of other senses like sound, vision, or lateral-line can be more ambiguous. Both left and right sensors can suggest a threat or obstacle to avoid. The system needs a WTA (winner-take-all) circuit.
In the diagram below a sound to the left drives a swimming muscle contraction on the right (‘C-bend’) followed by bilateral swimming and the reverse for a sound to the right. In the R4.mc circuit, the crossing is downstream of the R4.mc neuron.

As a first attempt at this WTA circuit, consider FFI (feedforward inhibition). A left sound drives the right choice and suppresses the left choice using an inhibitory GABA interneuron, and the flipped circuit for a right sound. The left diagram above shows this circuit. When one side significantly outweighs the other, the WTA circuit works by suppressing the other side, and we have the basis of a decision circuit.
However, this circuit runs into trouble when both sides are nearly equal or are equal. Because of the mutual inhibition, the choice neurons may not register any choice at all [Koyama et al 2016]. In some situations like avoiding an obstacle, stopping may be a reasonable outcome, but continuing to move forward without turning is not. In other cases like escaping a predator, not acting can be fatal. What we need is a true WTA circuit, not a circuit that fails for nearly equal input.
One solution is to add disinhibition to the FFI [Koyama et al 2016]. The diagram on the right shows the addition of FFI disinhibition. Each inhibitory neuron also inhibits its opposing peer with the effect of disinhibiting its primary sensor. The Koyama study examined these FFI circuits with and without disinhibition, showing that the disinhibition circuit sharpens the choice, making a better WTA circuit with a minimal zone of indecision.
Habenula and R.ip in R1
The R1 area near the MHB also includes Hb (Habenula) input to the hindbrain, projecting to both R1.ip (interpeduncular nucleus) and V.dr (dorsal raphe), both derived from ventral R1, and to V.mr (median raphe) derived from ventral R2-R3. Hb is the main input to R1.ip. Hb is derived form the M.pineal area and is involved with phototaxis [Chen X and Engert 2014], chemotaxis [Chen WY et al 2019], thermotaxis [Palieri 2023], and CO2 avoidance [Koide et al 2018], essentially a gradient-following system. The projections from R1.ip are less well understood, but R1.ip.r (rostral R1.ip) appears to be part of the head direction circuit [Petrucco et al 2023], and R1.ip.c (caudal R1.ip) is strongly left vs right directional [Dragomir et al 2020]. The location and function of R1.ip.c is similar to R2.artr (anterior rhombocephalon turning region), but no studies have explored whether they are connected areas or independent.
Unfortunately, the detailed action output path has not been detailed, although logically the Hb-R1.ip driven taxis behavior must connect with the locomotion motor somehow.
Filter feeding
The second major hindbrain system develops to support filter feeding and digestion. Since early aquatic vertebrates don’t need to distinguish filter feeding from breathing, a relatively simple system suffices. Adding filter feeding as an addition to locomotion introduces major decisions between the two systems, because the animal needs to decide when to filter feed and when to move.
While the locomotor hindbrain is similar to earlier chordate Amphioxus locomotion, the Amphioxus filter feeding differs from ascidian and vertebrate feeding because Amphioxus uses cilia for water flow and ascidian and early vertebrates actively pump water through pharyngeal arches [Li S and Wang 2021]. The ascidian filter feeding and digestion neurons are marked by the phox2 transcription factor [Gigante et al 2023]. In vertebrates, phox2 marks the visceral nervous system including branchial arch muscles like the face, jaw, neck, and pharynx, with corresponding hindbrain areas R.nst (nucleus of the solitary tract), N5 (trigeminal jaw), N7 (facial nerve), R.na (nucleus ambiguus), R.m10 (dorsal vagus motor), N10 [Dufour et al 2006].
Vertebrate hindbrain feeding and breathing is a two phase pumping system [Li S and Wang 2021]. The buccal cavity (mouth area) bounded by lips. In lamprey, the N5 muscles in R2-R3 expel water from the buccal cavity and rebound, passive recoil brings food and water into the mouth. A second phase drives the water through the pharyngeal slits, into a posterior operculum area, which is emptied by N7 muscles in R4-R5.

Amphioxus breathing is mostly through the skin and the pharyngeal slits are exclusively for filter feeding. The muscular filter feeding improved feeding, which opened a space to use the pharyngeal slits for a second purpose of breathing [Li S and Wang 2021]. The introduction of the jaw introduced new feeding opportunities beyond filter feeding, which adapted the older system to new requirements. Similarly, air breathing adapted some of the same circuits. The lamprey respiratory rhythm generator is in R2.m7 similar in location and function to the mammalian pre-Bötzinger area.
Filter feeding vs locomotion
Because sedentary filter feeding is mutually exclusive with locomotion, a sedentary filter feeder needs to stop filter feeding when moving away from a threat or because of low food yields, and start after arriving at a new food spot. Feeding can fail for several reasons, either from lack of available food, toxic food or digestion problems, or environmental hazards that trigger itch-like nociceptors, or possible predator threats. If sand becomes lodged in the pharyngeal slit filter, the animal can stop normal filter feeding and instead pump water in the opposite direction to clear the debris. For any of these problems, the animal needs to stop filter feeding, and often should move from the current location and find a new place to feed.
As part of the circuit for these decisions, the rostral R1.pb (parabrachial nucleus) is strongly tied to the more caudal R.nst (nucleus of the solitary tract) feeding and digestion areas. Unlike the areas discussed above, R1.pb is not primarily a rhythmic CPG center, although it is involved in breathing [Arthurs et al 2023]. It integrates a broad range of feeding, digestive, and visceral information, including taste, both positive like sweet and negative like bitter, and digestive signals like the detection of toxins in the gut. R1.pb also includes nociceptive signals from both N5 and N.sp, including itch. R1.pb CGRP (an alarm peptide) controls meal termination [Campos et al 2016] and signals danger [Campos et al 2018], essentially acting like a general alarm [Palmiter 2018], and suppressing feeding from threat [Yang et al 2021].
Other R1 areas are also involved in a decision to stop filter feeding, and move to avoid the area. V.lc (locus coerulus source of noradrenaline), also in R1, is a phox2a region that can suppress feeding and encourage place avoidance [Yang et al 2021]. Similarly, the serotonin V.mr (median raphe) from R2-R3, mentioned above in connection with Hb, is also associated with avoidance.
The point for this essay is to show that the hindbrain includes higher-level decision making between two systems as well as decision making within hindbrain systems [Cisek 2022].
Sleep
The hindbrain is also associated with sleep. The area around R1.pb and V.lc is strongly associated with waking [Ao et al 2021] and is one of the only areas where a single lesion can produce a coma [Fuller et al 2011]. One the sleep-promoting side, an area near N7 (R5-R6) region, named R5.pf (prefacial) is associated with sleep [Anaclet et al 2014], [Anaclet and Fuller 2017], [Chen MC et al 2020]. The two regions are interconnected with R5.pf suppressing the wake-generating region in R1.
Vestibular locomotion
Unlike crawling on the sea floor, vertebrate swimming is three dimensional, which greatly complicates stability, especially considering external forces, currents, and wave motion. Pre-vertebrate chordates like the ascidian tadpole swim in a helical pattern [McHenry and Strother 2003], which provides some swimming stability, but impairs the directionality of any sensing and navigation system. Amphibian tadpoles also swim in a spiral pattern [Jamieson and Roberts 2000], but adult swimming is stabilized as their vestibular and visual systems develop.
The vestibular system of early zebrafish larva is simplified with only one of the otoliths (the utricle) and none of the semicircular canals have developed [Bianco 2012]. Early development suggests pitch and roll posture correction near R4 [Straka and Baker 2013]. Still, this one system is critical for the larva’s survival and corrects errors in pitch, allowing the larva to consistently choose up or down using R4.tan (tangential nucleus near R4) [Straka and Baker 2013]. These corrections are a combination of the vestibular system in the hindbrain and optic flow from M.pt (pretectum), which are relayed to pitch-specific posture systems in the hindbrain [Wang K et al 2019]. Because the otolith can’t distinguish forward acceleration from the head tilting backward, the optic flow is critical for distinguishing the forward movement until the semicircular canals develop [Bianco et al 2012].

Vestibular information uses N8 (vestibuloauditory nerve), which arrives at R4, near the R4.mc Mauthner acoustic escape cells. Vestibular information is split according to direction (pitch, roll, yaw) into specialized nuclei, which covers almost the entire hindbrain [Straka and Baker 2013]. The lamprey divisions include rostral R.aon and caudal R.pon for semicircular canal processing and an intermediary R.ion for otolith processing. These areas are also interconnected, providing more complex capabilities than simple reflexes. The vestibular system is organized by motor output, not by vestibular input [Straka and Baker 2013]. So, the pitch area receives vestibular input from N8 and optic flow from M.pt and combines them to produce corrective swimming. The information from optic flow to the vestibular system uses mlf (medial longitudinal fasciculus).
To control swimming, the lamprey’s swimming systems are organized into four columns along the caudal hindbrain. Different combinations of the columns provide different corrective motions such as rolling left or right, or pitching up or down [Deliagina et al 2014]. The system is organized by motor output, so rolling to the right is handled by a specific R.rs group. 68% of R.rs that respond to vestibular input respond only in one plane (yaw, pitch, roll), and 25% respond to rotation in more than one plane. This system is handled by the hindbrain. With a unilateral vestibular lesion, the lamprey continuously rolls [Deliagina and Fegerstedt 2000]. Lamprey swimming can be continuously modulated to control direction, speed, and posture (pitch and roll) [Sankrithi and O’Malley 2010].
Note that non-image-forming photoreceptors can help posture and stability, even without optic flow. Like the simple phototaxis of zooplankton [Randel and Jékely 2016], when the lamprey is rolled to one side, the photoreceptor on the lower side will report less light than the one pointing toward the surface, and the posture system can use this different to roll the animal to match the light from both eyes. In fish this is called the dorsal righting reflex.
Although the ascidian tadpole has a gravity-sensing otolith, which is used for geotaxis, this otolith is not homologous to the vertebrate vestibular system.
Cerebellum-like regions
The vestibular senses have an immediate issue that the animal’s own motion produces vestibular signals, which would cause problems for posture control such as pitch adaptation. The larva zebrafish is a good example because only the otolith utricle part of the vestibular system is available, but it can’t distinguish forward acceleration from pitch tilting [Bianco et al 2012]. A naive posture correction might drive the animal up or down simply because the animal moved forward. As an improvement the system needs to take the animal’s own movement into account, which is a function of the cerebellum in R1.
The cerebellum and cerebellum-like structures are adaptive filters that learn how the animal’s own movement affects sensory input, including the lateral line and vestibular system [Bell et al 2008], [Montgomery et al 2012].
The cerebellum and cerebellum-like systems are too complex to cover in detail, but they’re important to mention because they’re connected with all the systems in the hindbrain.
Vestibular optic stabilization
The hindbrain also includes stabilization for the image-forming eye, both using VOR (vestibular-optomotor reflex), where vestibular data stabilizes the visual image using eye movements, OKR (optokinetic reflex), where optic flow stabilizes the image using eye movements, and OMR (optomotor reflex), where optic flow drives body movements to stabilize the image. OKR and OMR are widely studied reflexes.
However, since these image-stabilization reflexes assume the animal has already evolved an image-forming eye that uses stable images, these reflexes must have developed after the eye. The image-forming eye drives the evolution of muscles to stabilize the image. But note that stability is not absolutely required for vision because optic flow can provide information even with a fixed eye.
This late development may explain the apparent disorganization of the eye movement system. Horizontal eye movement in one direction uses motor neurons in the caudal hindbrain N6 (abducens) near N5, but the opposing movement is in the midbrain N3 (oculomotor nerve) near the OT and M.pt.

In the larval zebrafish the semicircular canals are not available, relying only on the utricle otolith, but that input for pitch VOR is ambiguous, because forward motion and pitch rotation produce the same information. Optic flow information from M.pt can disambiguate between the two, and eye stabilization uses a combination of vestibular and optical information [Bianco et al 2012]. Roll stabilization also uses vestibular and optical data.
Saccades
The hindbrain generates spontaneous saccades with the generator in ventral R2-R3 [Ramirez and Aksay 2021], which may correspond to R2.artr [Leyden et al 2021]. Saccades intersperse with fixation [Ramirez and Aksay 2021], where the eye maintains a consistent remembered position. 19% of spontaneous active hindbrain are saccade related. Although the OT is studied as a saccade center, the hindbrain can generate saccades on its own, although it can’t target them like the OT can.
R.vpni eye position integrator
The target eye position is managed by R.vnpi (vestibular position integrator) in the hindbrain near R6 that maintains the fixation eye position as the animal moves, updated by VOR or OKR [Aksay et al 2007], [Gonçalves et al 2014], [Lee MM et al 2015], [Miri et al 2011]. The velocity to position integrator has a long persistent time decay between 10s and 100s. It’s used as an example of a recurrent line attractor [Seung 1996], but the cells of R.vpni have multiple persistence times across neurons, varying from 2.5s to 14s [Miri et al 2011], contrary to the precise homogeneity of the canonical line attractor.
Hindbrain summary
The point of this essay is to demonstrate the hindbrain as an evolution of successive capabilities and to show how it is self-sufficient to control its own actions. Its action selection is determined as a hindbrain circuits with feedforward and feedback inhibition and even making decisions by integrate-to-threshold processes similar to the cortex and OT. As the [Borisyuk et al 2017] study suggests, the selection of the action paths can be generated from a simple neural model. In particular these action selections do not require midbrain or forebrain systems like the OT or basal ganglia.
[Humphries et al 2007] make a similar argument with the hindbrain “reticular formation,” pointing out that basal ganglia lesions do not completely impair behavior, even if the animal is decerebrate, where all of the brain anterior to the OT is removed, although the hindbrain-only behavior is limited. They point out that both descending and ascending information is available as an electrical bus like system, where each cluster can sample and send information to and from the bus. The more detailed tadpole studies described here with R.din/chx10 receives widespread inputs [Huang et al 2013] and R.ain provides widespread lateral feedback inhibition, while R.cin provides contralateral inhibition and R.din clusters provide recurrent excitation.
Despite the older “reticular” description, the hindbrain has a strongly modular hierarchical organization [Vishwanathan et al 2024]. For posture control, the lamprey has four organized motoneuron pools that fire differently to manage forward swimming, turns, pitch, and roll [Zelenin et al 2001]. As covered above, the hindbrain is highly structured both by its segmentation into functionally distinct rhombomeres and columns. The hindbrain is arranged into stripes of broad neuron classes (glutamate, glycine, gaba, serotonin), and within regions neurons are stacked according to age [Koyama et al 2011]. The genetic stripes of alternating excitatory (glutamate) and inhibitory (gaba and glycine) stripes [Soffe et al 2009], [Feierstein et al 2023], [Severi et al 2018] with the ventral locomotor marked by chx10 [Agha et al 2024].
The major hindbrain division of locomotor vs feeding/breathing is marked by different transcription factors. The ascidian-derived filter feeding system is marked by phox2b, while locomotor systems are typified by lhx3 [Mazzoni et al 2023].
The point isn’t to say that the hindbrain is independent of the rest of the brain. It’s more that the rest of the brain builds on basic decision making from the hindbrain. Now, an interesting detail from [Humphries et al 2007] is that they contrast the hindbrain to a decerebrate lesion anterior to OT, not the intact animal to a MHB (midbrain-hindbrain boundary) lesion. So, their argument leaves open the interesting question of the caudal midbrain.
Missing
This essay doesn’t explain Ppt (pedunculopontine tegmental), P.ldt (laterodorsal tegmental), S.nr (substantia nigra pars reticulata), V.rn (serotonin raphe nuclei), V.rmtg (gaba inhibition of Vta), and Vta.g (also gaba inhibition of Vta), all of which derive from the hindbrain, specifically R1. Most of these are important components of the basal ganglia, which has critical components in the forebrain and midbrain.
Midbrain additions to hindbrain functions
Understanding how the hindbrain works by itself can provide suggestions for understanding other areas like the midbrain. [Larbi et al 2022] studied the trunk touch escape swimming, focusing on MHB lesions disconnecting the hindbrain from the midbrain. Lesioning the midbrain did not eliminate the trunk escape, but it did slow the response and impair the WTA left vs right decision. With the MHB lesion, more touches produces simultaneous left and right premotor drives, which slowed the response by conflict. When the midbrain was allowed to participate, more decisions were clearly left or right. The midbrain then improves decision-making accuracy, even if it’s not absolutely required.
Similarly, without the midbrain the tadpole stopped swimming 23s after trunk stimulation, but with the midbrain the tadpole stopped swimming in 3s. The trunk to M.dmd connections is not sufficient to explain the results [Larbi et al 2022]. This suggests that the stopping decision has important components in the midbrain and/or forebrain.
Knowledge of the hindbrain to improve study of the midbrain. The midbrain could be studied as driving the hindbrain, with the hindbrain as motor output, or the midbrain could be studied as modulating existing hindbrain capabilities. The difference between these approaches might simplify understanding of the midbrain.
References
Agha MA, Kishore S, McLean DL. Cell-type-specific origins of locomotor rhythmicity at different speeds in larval zebrafish. Elife. 2024 Sep 17;13:RP94349.
Aksay E, Olasagasti I, Mensh BD, Baker R, Goldman MS, Tank DW. Functional dissection of circuitry in a neural integrator. Nat Neurosci. 2007 Apr;10(4):494-504.
Anaclet C, Ferrari L, Arrigoni E, Bass CE, Saper CB, Lu J, Fuller PM. The GABAergic parafacial zone is a medullary slow wave sleep-promoting center. Nat Neurosci. 2014 Sep;17(9):1217-24.
Anaclet C, Fuller PM. Brainstem regulation of slow-wave-sleep. Curr Opin Neurobiol. 2017 Jun;44:139-143. doi: 10.1016/j.conb.2017.04.004.
Ao Y, Yang B, Zhang C, Wu B, Zhang X, Xing D, Xu H. Locus Coeruleus to Paraventricular Thalamus Projections Facilitate Emergence From Isoflurane Anesthesia in Mice. Front Pharmacol. 2021 Apr 27;12:643172.
Arthurs JW, Pauli JL, Palmiter RD. Activation of Parabrachial Tachykinin 1 Neurons Counteracts Some Behaviors Mediated by Parabrachial Calcitonin Gene-related Peptide Neurons. Neuroscience. 2023 May 1;517:105-116.
Athira A, Dondorp D, Rudolf J, Peytral O, Chatzigeorgiou M. Comprehensive analysis of locomotion dynamics in the protochordate Ciona intestinalis reveals how neuromodulators flexibly shape its behavioral repertoire. PLoS Biol. 2022 Aug 4;20(8):e3001744.
Bell, Curtis C., Victor Han, and Nathaniel B. Sawtell. Cerebellum-like structures and their implications for cerebellar function. Annu. Rev. Neurosci. 31 (2008): 1-24.
Bhattacharyya K, McLean DL, MacIver MA. Visual Threat Assessment and Reticulospinal Encoding of Calibrated Responses in Larval Zebrafish. Curr Biol. 2017 Sep 25;27(18):2751-2762.e6.
Bianco IH, Ma LH, Schoppik D, Robson DN, Orger MB, Beck JC, Li JM, Schier AF, Engert F, Baker R. The tangential nucleus controls a gravito-inertial vestibulo-ocular reflex. Curr Biol. 2012 Jul 24;22(14):1285-95.
Bouvier J, Caggiano V, Leiras R, Caldeira V, Bellardita C, Balueva K, Fuchs A, Kiehn O. Descending Command Neurons in the Brainstem that Halt Locomotion. Cell. 2015 Nov 19;163(5):1191-1203.
Borisyuk R, Merrison-Hort R, Soffe SR, Koutsikou S, Li WC. To swim or not to swim: A population-level model of Xenopus tadpole decision making and locomotor behaviour. Biosystems. 2017 Nov;161:3-14.
Bostwick M, Smith EL, Borba C, Newman-Smith E, Guleria I, Kourakis MJ, Smith WC. Antagonistic Inhibitory Circuits Integrate Visual and Gravitactic Behaviors. Curr Biol. 2020 Feb 24;30(4):600-609.e2.
Brocard F, Ryczko D, Fénelon K, Hatem R, Gonzales D, Auclair F, Dubuc R. The transformation of a unilateral locomotor command into a symmetrical bilateral activation in the brainstem. J Neurosci. 2010 Jan 13;30(2):523-33.
Bubnys A, Kandel H, Kao LM, Pfaff D, Tabansky I. Hindbrain V2a Neurons Pattern Rhythmic Activity of Motor Neurons in a Reticulospinal Coculture. Front Neurosci. 2019 Oct 17;13:1077.
Buhl E, Roberts A, Soffe SR. The role of a trigeminal sensory nucleus in the initiation of locomotion. J Physiol. 2012 May 15;590(10):2453-69.
Buhl E., Soffe S.R., Roberts A. Sensory initiation of a co-ordinated motor response: synaptic excitation underlying simple decision-making. J. Physiol. 2015;593:4423–4437.
Campos CA, Bowen AJ, Schwartz MW, Palmiter RD. Parabrachial CGRP Neurons Control Meal Termination. Cell Metab. 2016 May 10;23(5):811-20.
Campos CA, Bowen AJ, Han S, Wisse BE, Palmiter RD, Schwartz MW. Cancer-induced anorexia and malaise are mediated by CGRP neurons in the parabrachial nucleus. Nat Neurosci. 2017 Jul;20(7):934-942.
Cangiano L, Grillner S. Commentary: Elimination of Left-Right Reciprocal Coupling in the Adult Lamprey Spinal Cord Abolishes the Generation of Locomotor Activity. Front Neural Circuits. 2018 Apr 25;12:34.
Cao C, Lemaire LA, Wang W, Yoon PH, Choi YA, Parsons LR, Matese JC, Wang W, Levine M, Chen K. Comprehensive single-cell transcriptome lineages of a proto-vertebrate. Nature. 2019 Jul;571(7765):349-354.
Chen MC, Sorooshyari SK, Lin JS, Lu J. A Layered Control Architecture of Sleep and Arousal. Front Comput Neurosci. 2020 Feb 12;14:8.
Chen WY, Peng XL, Deng QS, Chen MJ, Du JL, Zhang BB. Role of Olfactorily Responsive Neurons in the Right Dorsal Habenula-Ventral Interpeduncular Nucleus Pathway in Food-Seeking Behaviors of Larval Zebrafish. Neuroscience. 2019 Apr 15;404:259-267.
Chen X, Engert F. Navigational strategies underlying phototaxis in larval zebrafish. Front Syst Neurosci. 2014 Mar 25;8:39.
Cisek P. Evolution of behavioural control from chordates to primates. Philos Trans R Soc Lond B Biol Sci. 2022 Feb 14
Cregg JM, Leiras R, Montalant A, Wanken P, Wickersham IR, Kiehn O. Brainstem neurons that command mammalian locomotor asymmetries. Nat Neurosci. 2020 Jun;23(6):730-740.
Currie SP, Doherty GH, Sillar KT. Deep-brain photoreception links luminance detection to motor output in Xenopus frog tadpoles. Proc Natl Acad Sci U S A. 2016 May 24;113(21):6053-8.
Dale N. Delayed production of adenosine underlies temporal modulation of swimming in frog embryo. J Physiol. 1998 Aug 15;511 ( Pt 1)(Pt 1):265-72.
Dale N. Resetting Intrinsic Purinergic Modulation of Neural Activity: An Associative Mechanism? The Journal of neuroscience : the official journal of the Society for Neuroscience. 2002;22:10461–10469.
Deliagina TG, Fagerstedt P. Responses of reticulospinal neurons in intact lamprey to vestibular and visual inputs. J Neurophysiol. 2000 Feb;83(2):864-78.
Deliagina TG, Beloozerova IN, Orlovsky GN, Zelenin PV. Contribution of supraspinal systems to generation of automatic postural responses. Front Integr Neurosci. 2014 Oct 1;8:76.
Dragomir EI, Štih V, Portugues R. Evidence accumulation during a sensorimotor decision task revealed by whole-brain imaging. Nat Neurosci. 2020 Jan;23(1):85-93.
Dufour HD, Chettouh Z, Deyts C, de Rosa R, Goridis C, Joly JS, Brunet JF. Precraniate origin of cranial motoneurons. Proc Natl Acad Sci U S A. 2006 Jun 6;103(23):8727-32.
Feierstein CE, de Goeij MHM, Ostrovsky AD, Laborde A, Portugues R, Orger MB, Machens CK. Dimensionality reduction reveals separate translation and rotation populations in the zebrafish hindbrain. Curr Biol. 2023 Sep 25;33(18):3911-3925.e6.
Ferrario A, Palyanov A, Koutsikou S, Li W, Soffe S, Roberts A, Borisyuk R. From decision to action: Detailed modelling of frog tadpoles reveals neuronal mechanisms of decision-making and reproduces unpredictable swimming movements in response to sensory signals. PLoS Comput Biol. 2021 Dec 13;17(12):e1009654.
Fuller PM, Sherman D, Pedersen NP, Saper CB, Lu J. Reassessment of the structural basis of the ascending arousal system. J Comp Neurol. 2011 Apr 1;519(5):933-56.
Gigante ED, Piekarz KM, Gurgis A, Cohen L, Razy-Krajka F, Popsuj S, Ali HS, Sundaram SM, Stolfi A. Specification and survival of post-metamorphic branchiomeric neurons in the hindbrain of a non-vertebrate chordate. bioRxiv [Preprint]. 2023 Aug 14:2023.06.16.545305.
Girdhar K, Gruebele M, Chemla YR. The Behavioral Space of Zebrafish Locomotion and Its Neural Network Analog. PLoS One. 2015 Jul 1;10(7):e0128668.
Gonçalves PJ, Arrenberg AB, Hablitzel B, Baier H, Machens CK. Optogenetic perturbations reveal the dynamics of an oculomotor integrator. Front Neural Circuits. 2014 Feb 28;8:10.
Hoyer J, Kolar K, Athira A, van den Burgh M, Dondorp D, Liang Z, Chatzigeorgiou M. Polymodal sensory perception drives settlement and metamorphosis of Ciona larvae. Curr Biol. 2024 Mar 25;34(6):1168-1182.e7. doi: 10.1016/j.cub.2024.01.041. Epub 2024 Feb 8. PMID: 38335959.
Huang KH, Ahrens MB, Dunn TW, Engert F. Spinal projection neurons control turning behaviors in zebrafish. Curr Biol. 2013 Aug 19;23(16):1566-73.
Humphries MD, Gurney K, Prescott TJ. Is there a brainstem substrate for action selection? Philos Trans R Soc Lond B Biol Sci. 2007 Sep 29;362(1485):1627-39.
Jamieson D, Roberts A. Responses of young Xenopus laevis tadpoles to light dimming: possible roles for the pineal eye. J Exp Biol. 2000 Jun;203(Pt 12):1857-67.
Johnson CJ, Razy-Krajka F, Zeng F, Piekarz KM, Biliya S, Rothbächer U, Stolfi A. Specification of distinct cell types in a sensory-adhesive organ important for metamorphosis in tunicate larvae. PLoS Biol. 2024 Mar 13;22(3):e3002555.
Kohashi T, Nakata N, Oda Y. Effective sensory modality activating an escape triggering neuron switches during early development in zebrafish. J Neurosci. 2012 Apr 25;32(17):5810-20.
Koide T, Yabuki Y, Yoshihara Y. Terminal Nerve GnRH3 Neurons Mediate Slow Avoidance of Carbon Dioxide in Larval Zebrafish. Cell Rep. 2018 Jan 30;22(5):1115-1123.
Koutsikou S, Merrison-Hort R, Buhl E, Ferrario A, Li WC, Borisyuk R, Soffe SR, Roberts A. A simple decision to move in response to touch reveals basic sensory memory and mechanisms for variable response times. J Physiol. 2018 Dec;596(24):6219-6233.
Koyama M, Kinkhabwala A, Satou C, Higashijima S, Fetcho J. Mapping a sensory-motor network onto a structural and functional ground plan in the hindbrain. Proc Natl Acad Sci U S A. 2011 Jan 18;108(3):1170-5.
Lacalli, T., & Candiani, S. (2017). Locomotory control in amphioxus larvae: new insights from neurotransmitter data. EvoDevo, 8, 1-8.
Larbi MC, Messa G, Jalal H, Koutsikou S. An early midbrain sensorimotor pathway is involved in the timely initiation and direction of swimming in the hatchling Xenopus laevis tadpole. Front Neural Circuits. 2022 Dec 21;16:1027831.
Lee MM, Arrenberg AB, Aksay ER. A structural and genotypic scaffold underlying temporal integration. J Neurosci. 2015 May 20;35(20):7903-20.
Leyden C, Brysch C, Arrenberg AB. A distributed saccade-associated network encodes high velocity conjugate and monocular eye movements in the zebrafish hindbrain. Sci Rep. 2021 Jun 16;11(1):12644.
Li S, Wang F. Vertebrate Evolution Conserves Hindbrain Circuits despite Diverse Feeding and Breathing Modes. eNeuro. 2021 Apr 28;8(2):ENEURO.0435-20.2021.
Li W.-C., Sautois B., Roberts A., Soffe S.R. Reconfiguration of a vertebrate motor network: specific neuron recruitment and context-dependent synaptic plasticity. J. Neurosci. 2007;27:12267–12276.
Li WC, Roberts A, Soffe SR. Specific brainstem neurons switch each other into pacemaker mode to drive movement by activating NMDA receptors. J Neurosci. 2010 Dec 8;30(49):16609-20.
Li WC, Soffe SR. Stimulation of Single, Possible CHX10 Hindbrain Neurons Turns Swimming On and Off in Young Xenopus Tadpoles. Front Cell Neurosci. 2019 Feb 18;13:47. doi: 10.3389/fncel.2019.00047.
Liu, B., Satou, Y. Foxg specifies sensory neurons in the anterior neural plate border of the ascidian embryo. Nat Commun 10, 4911 (2019).
Marquart GD, Tabor KM, Bergeron SA, Briggman KL, Burgess HA. Prepontine non-giant neurons drive flexible escape behavior in zebrafish. PLoS Biol. 2019 Oct 15;17(10):e3000480.
Martorell N, Medan V. Audiovisual integration in the Mauthner cell enhances escape probability and reduces response latency. Sci Rep. 2022 Jan 20;12(1):1097.
Mazzoni EO, Mahony S, Closser M, Morrison CA, Nedelec S, Williams DJ, An D, Gifford DK, Wichterle H. Synergistic binding of transcription factors to cell-specific enhancers programs motor neuron identity. Nat Neurosci. 2013 Sep;16(9):1219-27.
McClellan AD. Response: Commentary: Elimination of Left-Right Reciprocal Coupling in the Adult Lamprey Spinal Cord Abolishes the Generation of Locomotor Activity. Front Neural Circuits. 2018 Aug 2;12:62.
McCrea DA, Rybak IA. Organization of mammalian locomotor rhythm and pattern generation. Brain Res Rev. 2008 Jan;57(1):134-46.
McHenry, M., & Strother, J. (2003). The kinematics of phototaxis in larvae of the ascidian Aplidium constellatum. Marine Biology, 142, 173-184.
Miri A, Daie K, Arrenberg AB, Baier H, Aksay E, Tank DW. Spatial gradients and multidimensional dynamics in a neural integrator circuit. Nat Neurosci. 2011 Aug 21;14(9):1150-9.
Montgomery, John C., David Bodznick, and Kara E. Yopak. The cerebellum and cerebellum-like structures of cartilaginous fishes. Brain Behavior and Evolution 80.2 (2012): 152-165.
Moult PR, Cottrell GA, Li WC. Fast silencing reveals a lost role for reciprocal inhibition in locomotion. Neuron. 2013 Jan 9;77(1):129-40.
Nokhbatolfoghahai M, Downie JR. Larval cement gland of frogs: comparative development and morphology. J Morphol. 2005 Mar;263(3):270-83.
Olivo P, Palladino A, Ristoratore F, Spagnuolo A. Brain Sensory Organs of the Ascidian Ciona robusta: Structure, Function and Developmental Mechanisms. Front Cell Dev Biol. 2021 Sep 6;9:701779.
Palieri, V., 2023. Principles in Homeostatic Navigation (Doctoral dissertation, Technische Universität München).
Palmiter RD. The Parabrachial Nucleus: CGRP Neurons Function as a General Alarm. Trends Neurosci. 2018 May;41(5):280-293.
Perrins R, Walford A, Roberts A. Sensory activation and role of inhibitory reticulospinal neurons that stop swimming in hatchling frog tadpoles. J Neurosci. 2002 May 15;22(10):4229-40.
Petrucco L, Lavian H, Wu YK, Svara F, Štih V, Portugues R. Neural dynamics and architecture of the heading direction circuit in zebrafish. Nat Neurosci. 2023 May;26(5):765-773.
Pryor, G.S., 2014. Tadpole nutritional ecology and digestive physiology: implications for captive rearing of larval anurans. Zoo biology, 33(6), pp.502-507.
Ramirez AD, Aksay ERF. Ramp-to-threshold dynamics in a hindbrain population controls the timing of spontaneous saccades. Nat Commun. 2021 Jul 6;12(1):4145.
Randel N, Jékely G. Phototaxis and the origin of visual eyes. Philos Trans R Soc Lond B Biol Sci. 2016 Jan 5;371(1685):20150042.
Rétaux S, Pottin K. A question of homology for chordate adhesive organs. Commun Integr Biol. 2011 Jan;4(1):75-7. doi: 10.4161/cib.4.1.13926. PMID: 21509185; PMCID: PMC3073277.
Roberts A, Hill NA, Hicks R. Simple mechanisms organise orientation of escape swimming in embryos and hatchling tadpoles of Xenopus laevis. J Exp Biol. 2000 Jun;203(Pt 12):1869-85.
Roberts A., Li W.C., Soffe S.R. How neurons generate behavior in a hatchling amphibian tadpole: an outline. Front. Behav. Neurosci. 2010;4:16.
Roberts A., Conte D., Hull M., Merrison-Hort R., Azad A.K., Buhl E., Borisyuk R., Soffe S.R. Can simple rules control development of a pioneer vertebrate neuronal network generating behavior? J. Neurosci. 2014;34:608–662.
Ryan K, Lu Z, Meinertzhagen IA. The CNS connectome of a tadpole larva of Ciona intestinalis (L.) highlights sidedness in the brain of a chordate sibling. Elife. 2016 Dec 6;5:e16962.
Ryan K, Lu Z, Meinertzhagen IA. Circuit Homology between Decussating Pathways in the Ciona Larval CNS and the Vertebrate Startle-Response Pathway. Curr Biol. 2017 Mar 6;27(5):721-728.
Ryczko D, Knüsel J, Crespi A, Lamarque S, Mathou A, Ijspeert AJ, Cabelguen JM. Flexibility of the axial central pattern generator network for locomotion in the salamander. J Neurophysiol. 2015 Mar 15;113(6):1921-40.
Sankrithi NS, O’Malley DM. Activation of a multisensory, multifunctional nucleus in the zebrafish midbrain during diverse locomotor behaviors. Neuroscience. 2010 Mar 31;166(3):970-93.
Seung HS. How the brain keeps the eyes still. Proc Natl Acad Sci U S A. 1996 Nov 12;93(23):13339-44.
Severi KE, Böhm UL, Wyart C. Investigation of hindbrain activity during active locomotion reveals inhibitory neurons involved in sensorimotor processing. Sci Rep. 2018 Sep 11;8(1):13615.
Shoenhard H, Jain RA, Granato M. The calcium-sensing receptor (CaSR) regulates zebrafish sensorimotor decision making via a genetically defined cluster of hindbrain neurons. Cell Rep. 2022 Dec 6;41(10):111790.
Soffe S.R. Triggering and gating of motor responses by sensory stimulation: behavioural selection in xenopus embryos. Proc. R. Soc. Lond. B. 1991;1991(246):197–203.
Soffe SR, Roberts A, Li WC. Defining the excitatory neurons that drive the locomotor rhythm in a simple vertebrate: insights into the origin of reticulospinal control. J Physiol. 2009 Oct 15;587(Pt 20):4829-44.
Stephens GJ, Johnson-Kerner B, Bialek W, Ryu WS. Dimensionality and dynamics in the behavior of C. elegans. PLoS Comput Biol. 2008 Apr 25;4(4):e1000028.
Straka H, Baker R. Vestibular blueprint in early vertebrates. Front Neural Circuits. 2013 Nov 19;7:182.
Svensson E, Jeffreys H, Li WC. The modulation of two motor behaviors by persistent sodium currents in Xenopus laevis tadpoles. J Neurophysiol. 2017 Jul 1;118(1):121-130.
Vishwanathan A, Sood A, Wu J, Ramirez AD, Yang R, Kemnitz N, Ih D, Turner N, Lee K, Tartavull I, Silversmith WM, Jordan CS, David C, Bland D, Sterling A, Seung HS, Goldman MS, Aksay ERF; Eyewirers. Predicting modular functions and neural coding of behavior from a synaptic wiring diagram. Nat Neurosci. 2024 Dec;27(12):2443-2454.
Wang K, Hinz J, Haikala V, Reiff DF, Arrenberg AB. Selective processing of all rotational and translational optic flow directions in the zebrafish pretectum and tectum. BMC Biol. 2019 Mar 29;17(1):29.
Yang B, Sanches-Padilla J, Kondapalli J, Morison SL, Delpire E, Awatramani R, Surmeier DJ. Locus coeruleus anchors a trisynaptic circuit controlling fear-induced suppression of feeding. Neuron. 2021 Mar 3;109(5):823-838.e6.
Zelenin PV, Grillner S, Orlovsky GN, Deliagina TG. Heterogeneity of the population of command neurons in the lamprey. J Neurosci. 2001 Oct 1;21(19):7793-803.